ATP Hydrolysis Powers Post-translational Translocation of Some Secretory Proteins in Yeast
In most eukaryotes, secretory proteins enter the ER by cotranslational translocation. In yeast, however, some secretory proteins enter the ER lumen after translation has been completed. In such post-translational translocation, the translocating protein passes through the same Sec61 translocon that is used in cotranslational translocation. However, the SRP and SRP receptor are not involved in post-translational translocation, and in such cases a direct interaction between the translocon and the signal sequence of the completed protein appears to be sufficient for targeting to the ER membrane. In addition, the driving force for unidirectional translocation across the ER membrane is provided by an additional protein complex known as the Sec63 complex and a member of the Hsp70 family of molecular chaperones known as BiP (see Chapter 3 for further discussion of molecular chaperones). The tetrameric Sec63 complex is embedded in the ER membrane in the vicinity of the translocon, whereas BiP is within the ER lumen. Like other members of the Hsp70 family, BiP has a peptide-binding domain and an ATPase domain. These chaperones bind and stabilize unfolded or partially folded proteins (see Figure 3-17).
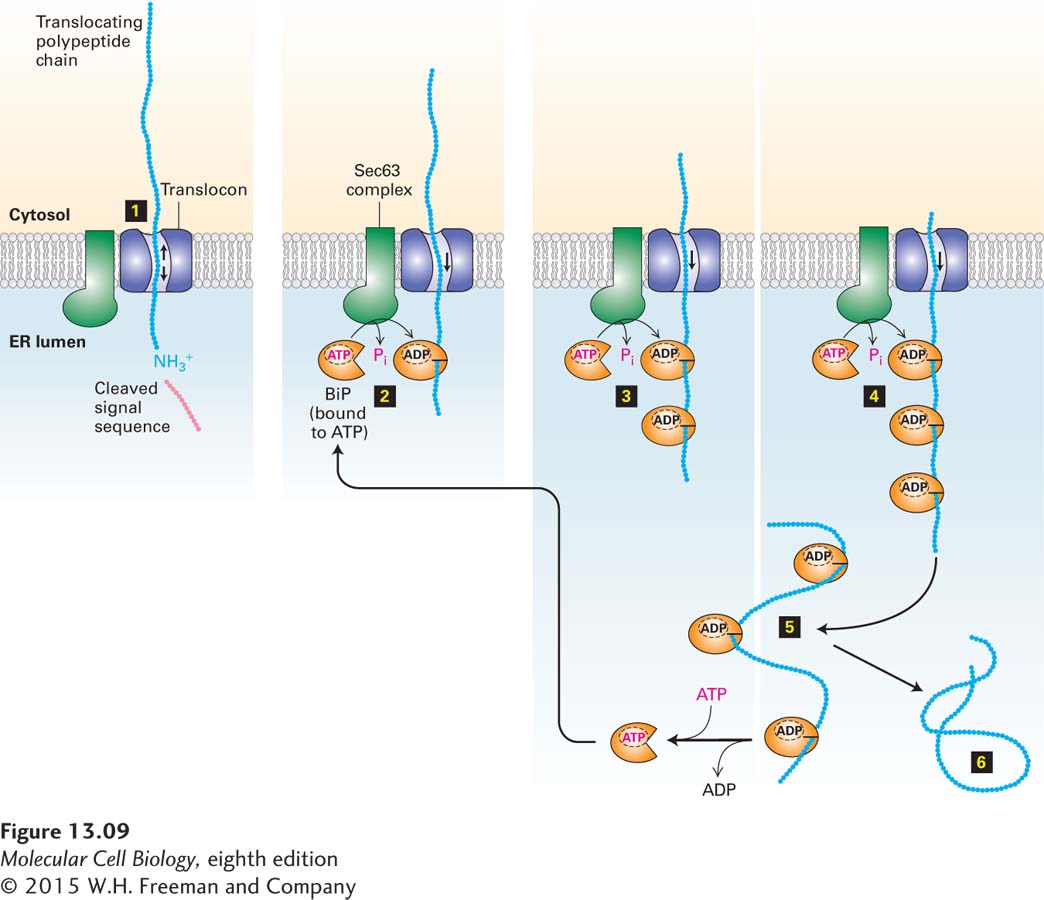
The current model for post-translational translocation of a protein into the ER is outlined in Figure 13-9. Once the N-terminal segment of the protein enters the ER lumen, signal peptidase cleaves the signal sequence just as in cotranslational translocation (step 1). Interaction of BiP·ATP with the luminal portion of the Sec63 complex causes hydrolysis of the bound ATP, producing a conformational change in BiP that promotes its binding to an exposed polypeptide chain (step 2). Since the Sec63 complex is located near the translocon, BiP is thus activated at sites where nascent polypeptides can enter the ER. Certain experiments suggest that, in the absence of binding to BiP, an unfolded polypeptide can freely slide back and forth within the translocon channel. Such random sliding motions rarely result in the entire polypeptide’s crossing the ER membrane. Binding of a molecule of BiP·ADP to the luminal portion of the polypeptide prevents backsliding of the polypeptide out of the ER. As further inward random sliding exposes more of the polypeptide on the luminal side of the ER membrane, successive binding of BiP·ADP molecules to the polypeptide chain acts as a ratchet, ultimately drawing the entire polypeptide into the ER within a few seconds (steps 3 and 4). On a slower time scale, the BiP molecules spontaneously exchange their bound ADP for ATP, leading to release of the polypeptide, which can then fold into its native conformation (steps 5 and 6). The recycled BiP·ATP is then ready for another interaction with Sec63. BiP and the Sec63 complex are also required for cotranslational translocation. The details of their role in this process are not well understood, but they are thought to act at an early stage of the process, such as the threading of the signal peptide into the pore of the translocon.
FIGURE 13-9Post-translational translocation. This mechanism is fairly common in yeast and probably occurs occasionally in higher eukaryotes. Small arrows inside the translocon represent random sliding of the translocating polypeptide inward and outward. Successive binding of BiP·ATP to entering segments of the polypeptide prevents the chain from sliding out toward the cytosol. See K. E. Matlack et al., 1997, Science277:938.
Page 592
The overall reaction carried out by BiP is an important example of how the chemical energy released by the hydrolysis of ATP can power the mechanical movement of a protein across a membrane. Bacterial cells also use an ATP-driven process for translocating completed proteins across the plasma membrane—in this case to be released from the cell. In bacteria, the driving force for translocation comes from a cytosolic ATPase known as the SecA protein. SecA binds to the cytoplasmic side of the translocon and hydrolyzes cytosolic ATP. By a mechanism that resembles the needle on a sewing machine, the SecA protein pushes segments of the polypeptide through the membrane in a mechanical cycle coupled to the hydrolysis of ATP.
As we will see, translocation of proteins across other eukaryotic organelle membranes, such as those of mitochondria and chloroplasts, also typically occurs by post-translational translocation. This explains why ribosomes are typically not found bound to these other organelles, as they are to the rough ER.