COPII Vesicles Mediate Transport from the ER to the Golgi
COPII vesicles were first recognized when cell-free extracts of yeast rough ER membranes were incubated with cytosol and a nonhydrolyzable analog of GTP. The vesicles that formed from the ER membranes had a distinct coat similar to that on COPI vesicles, but composed of different proteins, designated COPII proteins. Yeast cells with mutations in the genes for COPII proteins are class B sec mutants and accumulate proteins in the rough ER (see Figure 14-4). Analysis of such mutants has revealed a set of proteins required for formation of COPII vesicles, including the proteins that make up the COPII vesicle coat.
Page 646
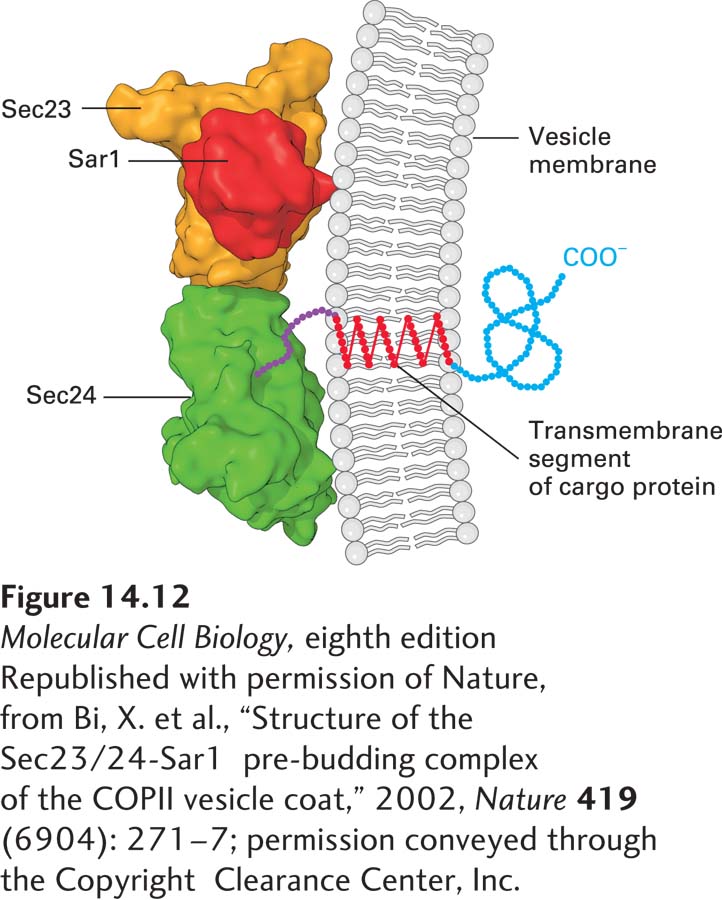
As described previously, formation of COPII vesicles is triggered when Sec12, a GEF in the ER membrane, catalyzes the exchange of bound GDP for GTP on cytosolic Sar1. This exchange induces binding of Sar1 to the ER membrane, followed by binding of a complex of Sec23 and Sec24 proteins (see Figure 14-8). The resulting ternary complex formed between Sar1·GTP, Sec23, and Sec24 is shown in Figure 14-12. This core coat protein complex then provides binding sites for the recruitment of a second complex of Sec13 and Sec31 proteins to complete the coat structure. Since pure Sec13 and Sec31 proteins can spontaneously assemble into cagelike lattices, it is thought that Sec13 and Sec31 form the structural scaffold for COPII vesicles. Finally, a large fibrous protein called Sec16, which is bound to the cytosolic surface of the ER, interacts with Sar1·GTP and the Sec13/31 and Sec23/24 complexes to organize the other coat proteins, increasing the efficiency of coat polymerization. Like Sec 13/31, clathrin has the ability to self-assemble into a coat-like structure, as will be discussed in Section 14.4.
[Republished with permission of Nature, from Bi, X. et al., “Structure of the Sec23/24-Sar1 pre-budding complex of the COPII vesicle coat,” 2002, Nature419(6904):271–7; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 14-12Three-dimensional structure of the ternary complex comprising the COPII coat proteins Sec23 and Sec24 and Sar1·GTP. Early in the formation of the COPII coat, Sec23 (orange)/Sec24 (green) complexes are recruited to the ER membrane by Sar1 (red) in its GTP-bound state. In order to form a stable ternary complex in solution for structural studies, the nonhydrolyzable GTP analog GppNHp is used. A cargo protein in the ER membrane can be recruited to COPII vesicles by the interaction of a tripeptide di-acidic sorting signal (purple) in the cargo protein’s cytosolic domain with Sec24. The likely positions of the COPII vesicle membrane and the transmembrane segment of the cargo protein are indicated. The N-terminal segment of Sar1 that tethers it to the membrane is not shown.
[Republished with permission of Nature, from Bi, X. et al., “Structure of the Sec23/24-Sar1 pre-budding complex of the COPII vesicle coat,” 2002, Nature419(6904):271–7; permission conveyed through the Copyright Clearance Center, Inc.]
Certain integral ER membrane proteins are specifically recruited into COPII vesicles for transport to the Golgi. The cytosolic segments of many of these proteins contain a di-acidic sorting signal (the key residues in this sequence are Asp-X-Glu, or DXE in the one-letter code) (see Table 14-2). This sorting signal, which binds to the Sec24 subunit of the COPII coat, is essential for the selective export of certain membrane proteins from the ER (see Figure 14-12). Biochemical and genetic studies have identified additional signals that help direct membrane cargo proteins into COPII vesicles. All of the known sorting signals bind to one or another site on the Sec24 subunit of COPII. Ongoing studies seek to determine how soluble cargo proteins are selectively loaded into COPII vesicles. For example, TANGO1 is an ER membrane protein that acts as a cargo receptor for collagen by simultaneously binding to collagen in the lumen and to the Sec24 subunit of the coat.
The inherited disease cystic fibrosis is characterized by an imbalance in chloride and sodium ion transport in the epithelial cells of the lungs, leading to fluid buildup and difficulty breathing. Cystic fibrosis is caused by mutations in a protein known as CFTR, which is synthesized as an integral membrane protein in the ER and is transported to the Golgi before being transported to the plasma membranes of epithelial cells, where it functions as a chloride channel. Researchers have recently shown that the CFTR protein contains a di-acidic sorting signal that binds to the Sec24 subunit of the COPII vesicle coat and is necessary for transport of the CFTR protein out of the ER. The most common CFTR mutation is a deletion of a phenylalanine at position 508 in the protein sequence (known as ΔF508). This mutation prevents normal transport of CFTR to the plasma membrane by blocking its packaging into COPII vesicles budding from the ER. Although the ΔF508 mutation is not in the vicinity of the di-acidic sorting signal, this mutation may change the conformation of the cytosolic portion of CFTR so that the signal is unable to bind to Sec24. Interestingly, a folded CFTR with this mutation will still function properly as a normal chloride channel. However, it never reaches the membrane; the disease state is therefore caused by the absence of the channel, rather than by a defective channel.
The experiments described previously in which the transit of VSVG-GFP in cultured mammalian cells was followed by fluorescence microscopy (see Figure 14-2) provided insight into the intermediates in ER-to-Golgi transport. In some cells, small fluorescent vesicles containing VSVG-GFP could be seen to form from the ER, move less than 1 µm, and then fuse directly with the cis-Golgi. In other cells, in which the ER was located several micrometers from the Golgi complex, several ER-derived vesicles were seen to fuse with one another shortly after their formation, forming what is termed the ER-to-Golgi intermediate compartment or the cis-Golgi network. These larger structures were then transported along microtubules to the cis-Golgi, much in the way vesicles in neurons are transported from the cell body, where they are formed, down the long axon to the axon terminus (see Chapter 18). Microtubules function like “railroad tracks,” enabling these large aggregates of transport vesicles to move long distances to their cis-Golgi destination. At the time the ER-to-Golgi intermediate compartment is formed, some COPI vesicles bud off from it, recycling some proteins back to the ER.