COPI Vesicles Mediate Retrograde Transport Within the Golgi and from the Golgi to the ER
COPI vesicles were first discovered when isolated Golgi fractions were incubated in a solution containing cytosol and a nonhydrolyzable analog of GTP (see Figure 14-9). Subsequent analysis of these vesicles showed that the coat is formed from large cytosolic complexes, called coatomers, composed of seven polypeptide subunits. Yeast cells containing temperature-sensitive mutations in COPI proteins accumulate proteins in the rough ER at the nonpermissive temperature and thus are categorized as class B sec mutants (see Figure 14-4). Although discovery of these mutants initially suggested that COPI vesicles mediate ER-to-Golgi transport, subsequent experiments showed that their main function is retrograde transport, both between Golgi cisternae and from the cis-Golgi to the rough ER (see Figure 14-11, right). Because COPI mutants cannot recycle key membrane proteins back to the rough ER, the ER gradually becomes depleted of ER proteins, such as v-SNAREs, that are necessary for COPII vesicle function. Eventually, vesicle formation from the rough ER grinds to a halt; secretory proteins continue to be synthesized but accumulate in the ER—the defining characteristic of class B sec mutants. The general ability of sec mutations involved in either COPI or COPII vesicle function to eventually block both anterograde and retrograde transport illustrates the fundamental interdependence of these two transport processes.
As discussed in Chapter 13, the ER contains several soluble proteins dedicated to the folding and modification of newly synthesized secretory proteins. They include the chaperone BiP and the enzyme protein disulfide isomerase, which are necessary for the ER to carry out its functions. Although such ER-resident luminal proteins are not specifically selected by COPII vesicles, their sheer abundance causes them to be continuously loaded passively into vesicles destined for the cis-Golgi. The transport of these soluble proteins back to the ER, mediated by COPI vesicles, prevents their eventual depletion.
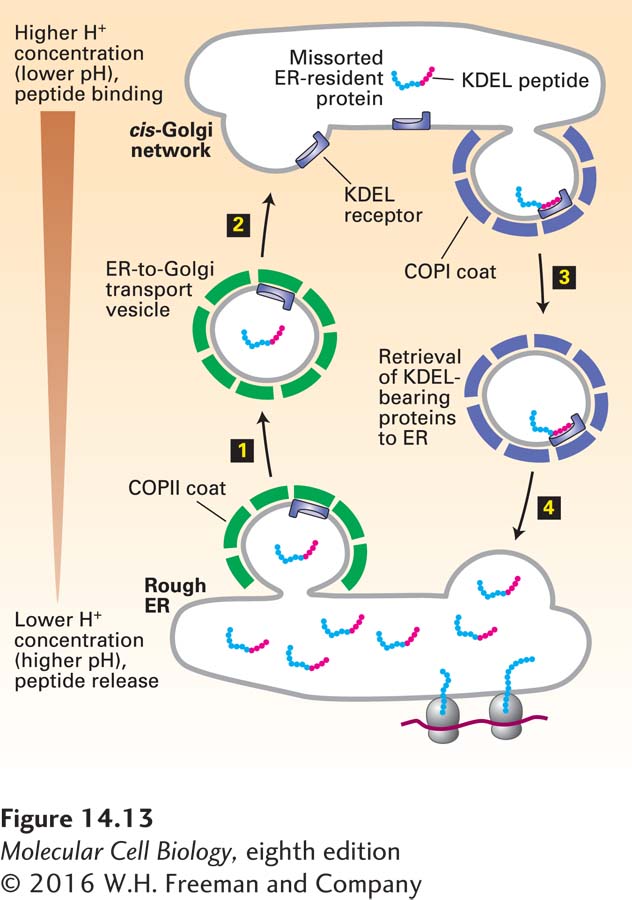
Most soluble ER-resident proteins carry a Lys-Asp-Glu-Leu (KDEL in the one-letter code) sequence at their C-terminus (see Table 14-2). Several experiments have demonstrated that this KDEL sorting signal is both necessary and sufficient to cause a protein bearing this sequence to be located in the ER. For instance, when a mutant protein disulfide isomerase lacking these four residues is synthesized in cultured fibroblasts, the protein is secreted. Moreover, if a protein that is normally secreted is altered so that it contains the KDEL sorting signal at its C-terminus, the protein is located in the ER. The KDEL sorting signal is recognized and bound by the KDEL receptor, a transmembrane protein found primarily on small transport vesicles shuttling between the ER and the cis-Golgi and on the cis-Golgi membrane. In addition, soluble ER-resident proteins that carry the KDEL signal have oligosaccharide chains bearing modifications that are catalyzed by enzymes found only in the cis-Golgi or cis-Golgi network; thus at some time these proteins must have left the ER and been transported at least as far as the cis-Golgi network. These findings indicate that the KDEL receptor acts mainly to retrieve soluble proteins containing the KDEL sorting signal that have escaped to the cis-Golgi network and return them to the ER (Figure 14-13). The KDEL receptor binds more tightly to its ligand at low pH, and it is thought that the receptor is able to bind KDEL peptides in the cis-Golgi and release these peptides in the ER because the pH of the Golgi is slightly lower than that of the ER.
FIGURE 14-13Role of the KDEL receptor in retrieval of ER-resident luminal proteins from the Golgi. ER luminal proteins, especially those present at high concentrations, can be passively incorporated into COPII vesicles and transported to the Golgi (steps 1 and 2). Many such proteins bear a C-terminal KDEL (Lys-Asp-Glu-Leu) sequence (red) that allows them to be retrieved. The KDEL receptor, located mainly in the cis-Golgi network and in both COPII and COPI vesicles, binds proteins bearing the KDEL sorting signal and returns them to the ER (steps 3 and 4). This retrieval system prevents depletion of ER luminal proteins such as those needed for proper folding of newly made secretory proteins. The binding affinity of the KDEL receptor is very sensitive to pH. The small difference between the pH of the ER and that of the Golgi favors binding of KDEL-bearing proteins to the receptor in Golgi-derived vesicles and their release in the ER. See J. Semenza et al., 1990, Cell61:1349.
Page 648
The KDEL receptor and other membrane proteins that are transported back to the ER from the Golgi contain a Lys-Lys-X-X sequence at the very end of their C-terminal segment, which faces the cytosol (see Table 14-2). This KKXX sorting signal, which binds to a complex of the COPI α and β subunits (two of the seven polypeptide subunits in the COPI coatomer), is both necessary and sufficient to incorporate membrane proteins into COPI vesicles for retrograde transport to the ER. Temperature-sensitive yeast mutants lacking COPIα or COPIβ are not only unable to bind the KKXX signal, but are also unable to transport proteins bearing this signal back to the ER, indicating that COPI vesicles mediate retrograde Golgi-to-ER transport.
A second sorting signal that targets proteins to COPI vesicles and thus enables recycling from the Golgi to the ER is a di-arginine sequence. Unlike the KKXX sorting signal, which must be located at the cytoplasmically oriented C-terminus of a protein, the di-arginine sorting signal can reside in any segment of a membrane protein that is on the cytoplasmic face of the membrane.
The partitioning of proteins between the ER and Golgi complex is a highly dynamic process depending on both COPII (anterograde) and COPI (retrograde) vesicles, with each type of vesicle responsible for recycling the components necessary for the function of the other type of vesicle. The organization of this partitioning process raises an interesting puzzle: how do vesicles preferentially use the v-SNAREs that specify fusion with the correct target membrane instead of the v-SNAREs that are being recycled and specify fusion with the donor membrane?
This basic question concerning correct membrane partitioning has recently been answered for COPII vesicles. After these vesicles form, the COPII coat proteins remain assembled long enough for the Sec23/Sec24 complex to interact with a specific tethering factor attached to the cis-Golgi membrane. Vesicle uncoating to expose the v-SNAREs is completed only after the COPII vesicle is already closely associated with the cis-Golgi membrane and the COPII v-SNAREs are in position to form complexes with their cognate t-SNAREs. Although COPII vesicles also carry COPI-specific v-SNARE proteins, which are being recycled back to the cis-Golgi, these COPI v-SNARE proteins never have the opportunity to form SNARE complexes with cognate ER-localized t-SNARE proteins.