Transport of a Protein Through the Secretory Pathway Can Be Assayed in Live Cells
The classic studies of G. Palade and his colleagues in the 1960s first established the order in which proteins move from one organelle to the next in the secretory pathway (see Classic Experiment 14-1). These early studies also showed that secretory proteins are never released into the cytosol—the first indication that transported proteins are always associated with some type of membrane-bounded intermediate. In these experiments, which combined pulse-chase labeling (see Figure 3-42) and autoradiography, radioactively labeled amino acids were injected into the pancreas of hamsters. At different times after injection, the animals were sacrificed and the pancreatic cells were immediately fixed with glutaraldehyde, sectioned, and subjected to autoradiography to visualize the locations of the radiolabeled proteins. Because the radioactive amino acids were administered in a short pulse, only those proteins synthesized immediately after injection were labeled, forming a distinct cohort of labeled proteins whose transport could be followed. In addition, because pancreatic acinar cells are dedicated secretory cells, almost all of the labeled amino acids in these cells were incorporated into secretory proteins, facilitating the observation of transported proteins.
Although autoradiography is rarely used today to localize proteins within cells, these early experiments illustrate the two basic requirements for any assay of intercompartmental transport. First, it is necessary to label a cohort of proteins in an early compartment so that their subsequent transfer to later compartments can be followed over time. Second, it is necessary to have a way to identify the compartment in which a labeled protein resides. Here we describe two modern experimental procedures for observing the intracellular trafficking of a secretory protein in almost any type of cell.
In both procedures, a gene encoding an abundant membrane glycoprotein (G protein) from vesicular stomatitis virus (VSV) is introduced into cultured mammalian cells either by transfection or simply by infecting the cells with the virus. The treated cells, even those that are not specialized for secretion, rapidly synthesize the VSV G protein on the ER as they would normal cellular secretory proteins. Use of a mutant gene encoding a temperature-sensitive VSV G protein allows researchers to turn subsequent transport of this protein on and off. At the restrictive temperature of 40 °C, newly made VSV G protein is misfolded and is therefore retained within the ER by the quality-control mechanisms discussed in Chapter 13, whereas at the permissive temperature of 32 °C, the protein is correctly folded and is transported through the secretory pathway to the cell surface. Importantly, the misfolding of the temperature-sensitive VSV G protein is reversible; thus when cells synthesizing mutant VSV G protein are grown at 40 °C and then shifted to 32 °C, the misfolded mutant VSV G protein that had accumulated in the ER will refold and be transported normally. This clever use of a temperature-sensitive mutation in effect defines a protein cohort whose subsequent transport can be followed.
In two variations of this basic procedure, transport of VSV G protein is monitored by different techniques. Studies using both of these modern trafficking assays came to the same conclusion as Palade’s early experiments: in mammalian cells, vesicle-mediated transport of a protein molecule from its site of synthesis on the rough ER to its arrival at the plasma membrane takes from 30 to 60 minutes.
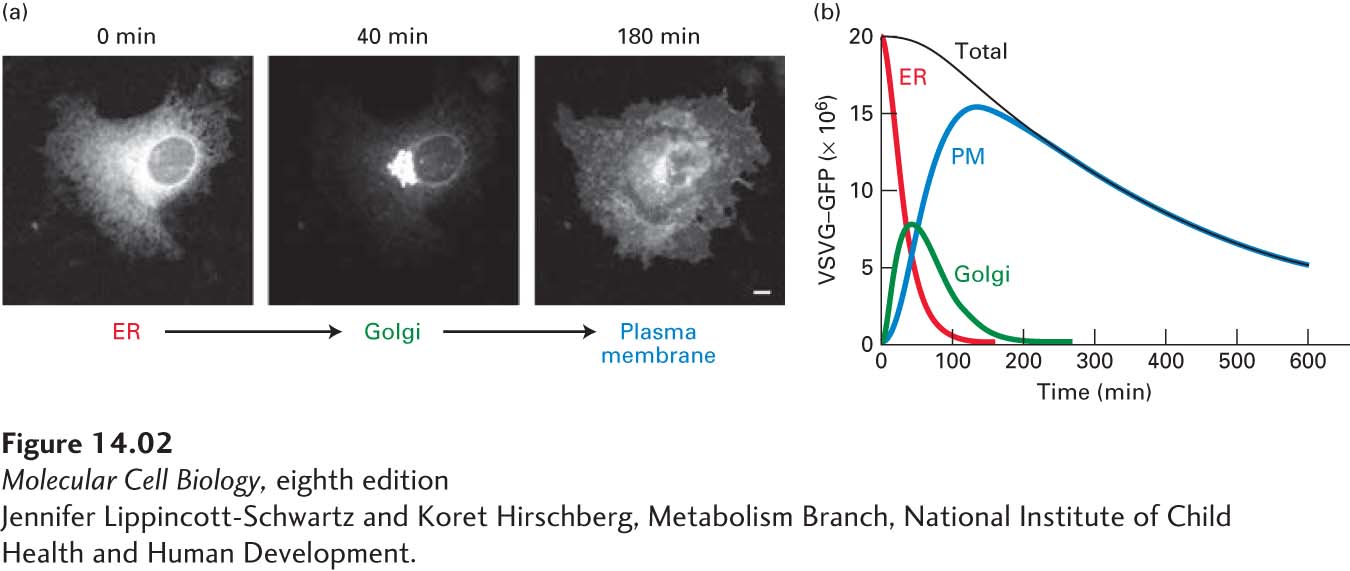
Microscopy of GFP-Labeled VSV G Protein One approach for observing the transport of VSV G protein employs a hybrid gene in which the viral gene is fused to the gene encoding green fluorescent protein (GFP), a naturally fluorescent protein (see Chapter 4). The hybrid gene is transfected into cultured cells by techniques described in Chapter 6. When cells expressing the temperature-sensitive form of the hybrid protein (VSVG-GFP) are grown at the restrictive temperature, VSVG-GFP accumulates in the ER, which appears as a lacy network of membranes when the cells are observed in a fluorescent microscope. When the cells are subsequently shifted to a permissive temperature, the VSVG-GFP can be seen to move first to the membranes of the Golgi complex, which are densely concentrated at the edge of the nucleus, and then to the cell surface (Figure 14-2a). By observing the distribution of VSVG-GFP at different times after shifting cells to the permissive temperature, researchers have determined how long VSVG-GFP resides in each organelle of the secretory pathway (Figure 14-2b).
Page 635
[Jennifer Lippincott-Schwartz and Koret Hirschberg, Metabolism Branch, National Institute of Child Health and Human Development.]
EXPERIMENTAL FIGURE 14-2Protein transport through the secretory pathway can be visualized by fluorescence microscopy of cells producing a GFP-tagged membrane protein. Cultured cells were transfected with a hybrid gene encoding the viral membrane glycoprotein VSV G linked to the gene for green fluorescent protein (GFP). A temperature-sensitive mutant version of the viral gene was used so that newly made hybrid protein (VSVG-GFP) was retained in the ER at 40 °C, but was released for transport at 32 °C. (a) Fluorescence micrographs of cells just before and at two times after they were shifted to the lower temperature. Movement of VSVG-GFP from the ER to the Golgi and finally to the cell surface occurred within 180 minutes. The scale bar is 5 µm. (b) Plot of the amount of VSVG-GFP in the endoplasmic reticulum (ER), Golgi, and plasma membrane (PM) at different times after the shift to the permissive temperature. The kinetics of transport from one organelle to another can be reconstructed from computer analysis of these data. The decrease in total fluorescence that occurs at later times probably results from slow inactivation of GFP fluorescence.
[Jennifer Lippincott-Schwartz and Koret Hirschberg, Metabolism Branch, National Institute of Child Health and Human Development.]
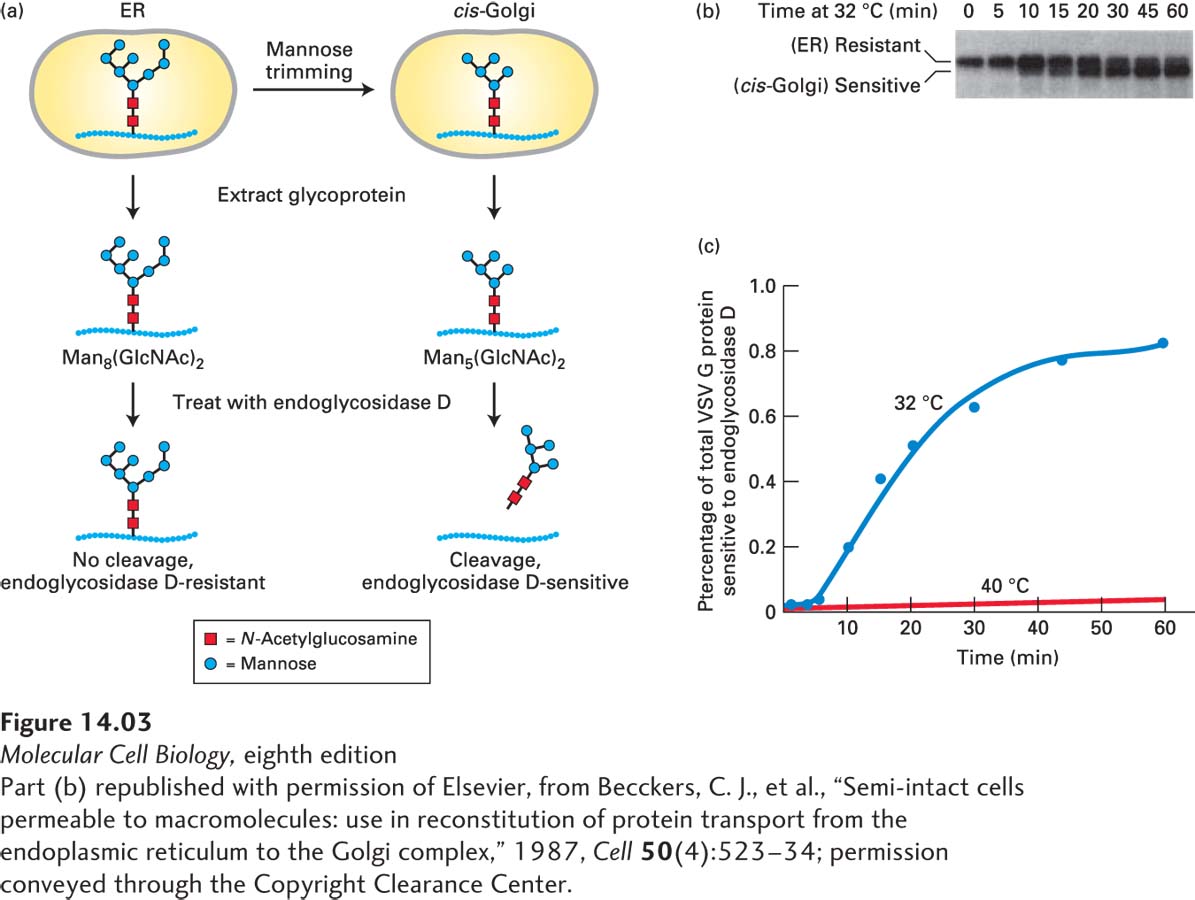
Detection of Compartment-Specific Oligosaccharide Modifications A second way to follow the transport of secretory proteins takes advantage of modifications to their carbohydrate side chains that occur at different stages of the secretory pathway. To understand this approach, recall that many secretory proteins leaving the ER are carrying one or more copies of the N-linked oligosaccharide Man8(GlcNAc)2, which are synthesized and attached to secretory proteins in the ER (see Figure 13-18). As a protein moves through the Golgi complex, different enzymes localized to the cis-, medial-, and trans-Golgi cisternae catalyze an ordered series of modifications to these core Man8(GlcNAc)2 chains, as discussed in a later section of this chapter. For instance, glycosidases that reside specifically in the cis-Golgi compartment sequentially trim mannose residues off the core oligosaccharide to yield a “trimmed” form, Man5(GlcNAc)2. Scientists can use a specialized carbohydrate-cleaving enzyme known as endoglycosidase D to distinguish glycosylated proteins that remain in the ER from those that have entered the cis-Golgi: trimmed cis-Golgi-specific oligosaccharides are cleaved from proteins by endoglycosidase D, whereas the core (untrimmed) oligosaccharide chains on secretory proteins within the ER are resistant to digestion by this enzyme (Figure 14-3a). Because a deglycosylated protein produced by endoglycosidase D digestion moves faster on an SDS gel than the corresponding glycosylated protein, these proteins can be readily distinguished (Figure 14-3b).
[Part (b) republished with permission of Elsevier, from Becckers, C. J., et al., “Semi-intact cells permeable to macromolecules: use in reconstitution of protein transport from the endoplasmic reticulum to the Golgi complex,” 1987, Cell50(4):523–34; permission conveyed through the Copyright Clearance Center.]
EXPERIMENTAL FIGURE 14-3Transport of a membrane glycoprotein from the ER to the Golgi can be assayed based on sensitivity to cleavage by endoglycosidase D. Cells expressing a temperature-sensitive VSV G protein were labeled with a pulse of radioactive amino acids at the nonpermissive temperature so that the labeled protein was retained in the ER. At periodic times after a return to the permissive temperature of 32 °C, VSV G protein was extracted from cells and digested with endoglycosidase D. (a) As proteins move to the cis-Golgi from the ER, the core oligosaccharide Man8(GlcNAc)2 is trimmed to Man5(GlcNAc)2 by enzymes that reside in the cis-Golgi compartment. Endoglycosidase D cleaves the oligosaccharide chains from proteins processed in the cis-Golgi, but not from proteins in the ER. (b) SDS-polyacrylamide gel electrophoresis of the digestion mixtures resolves the resistant, uncleaved (slower-migrating) and sensitive, cleaved (faster-migrating) forms of labeled VSV G protein. Initially, as this gel shows, all of the VSV G protein was resistant to digestion, but over time, an increasing fraction was sensitive to digestion, reflecting transport of the protein from the ER to the Golgi and its processing there. In control cells kept at 40 °C, only slow-moving, digestion-resistant VSV G protein was detected after 60 minutes (not shown). (c) A plot of the percentage of VSV G protein that is sensitive to digestion, derived from electrophoretic data, reveals the time course of ER-to-Golgi transport.
[Part (b) republished with permission of Elsevier, from Becckers, C. J., et al., “Semi-intact cells permeable to macromolecules: use in reconstitution of protein transport from the endoplasmic reticulum to the Golgi complex,” 1987, Cell50(4):523–34; permission conveyed through the Copyright Clearance Center.]
This type of assay can be used to track movement of VSV G protein in virus-infected cells pulse-labeled with radioactive amino acids. Immediately after labeling, all the labeled VSV G protein is still in the ER and, upon extraction, is resistant to digestion by endoglycosidase D, but over time, the fraction of the extracted glycoprotein that is sensitive to digestion increases. This conversion of VSV G protein from an endoglycosidase D–resistant form to an endoglycosidase D–sensitive form corresponds to vesicular transport of the protein from the ER to the cis-Golgi. Note that transport of VSV G protein from the ER to the Golgi takes about 30 minutes, as measured either by the assay based on oligosaccharide processing or by fluorescence microscopy of VSVG-GFP (Figure 14-3c). A variety of assays based on specific carbohydrate modifications that occur in later Golgi compartments have been developed to measure progression of VSV G protein through each stage of the Golgi complex.