The Acidic pH of Late Endosomes Causes Most Receptor-Ligand Complexes to Dissociate
The overall rate of endocytic internalization of the plasma membrane is quite high: cultured fibroblasts regularly internalize 50 percent of their cell-surface proteins and phospholipids each hour. Most cell-surface receptors that undergo endocytosis will repeatedly deposit their ligands within the cell and then recycle to the plasma membrane to mediate internalization of ligand molecules once again. The LDL receptor, for instance, makes one round trip into and out of the cell interior every 10–20 minutes, for a total of about a hundred trips in its 20-hour life span.
Internalized receptor-ligand complexes commonly follow the pathway depicted for the M6P receptor in Figure 14-22 and the LDL receptor in Figure 14-29. Endocytosed cell-surface receptors typically dissociate from their ligands within late endosomes, which appear as spherical vesicles with tubular branching membranes located a few micrometers from the cell surface. The original experiments that defined the late endosome as a sorting compartment used the asialoglycoprotein receptor. This liver-specific receptor mediates the binding and internalization of abnormal glycoproteins whose oligosaccharides terminate in galactose rather than the normal sialic acid; hence the name asialoglycoprotein. Electron microscopy of liver cells perfused with asialoglycoprotein reveal that 5–10 minutes after internalization, ligand molecules are found in the lumen of late endosomes, while the tubular membrane extensions are rich in receptor and rarely contain ligand. These findings indicate that the late endosome is the organelle in which receptors and ligands are uncoupled.
Page 663
The dissociation of receptor-ligand complexes in late endosomes occurs not only in the endocytic pathway, but also in the delivery of soluble lysosomal enzymes via the secretory pathway (see Figure 14-22). As discussed in Chapter 11, the membranes of late endosomes and lysosomes contain V-class proton pumps that act in concert with Cl− channels to acidify the vesicle lumen (see Figure 11-14). Most receptors, including the M6P receptor and cell-surface receptors for LDL and asialoglycoprotein, bind their ligands tightly at neutral pH, but release their ligands if the pH is lowered to 6.0 or below. The late endosome is the first organelle encountered by receptor-ligand complexes whose luminal pH is sufficiently acidic to promote dissociation of most endocytosed receptors from their tightly bound ligands.
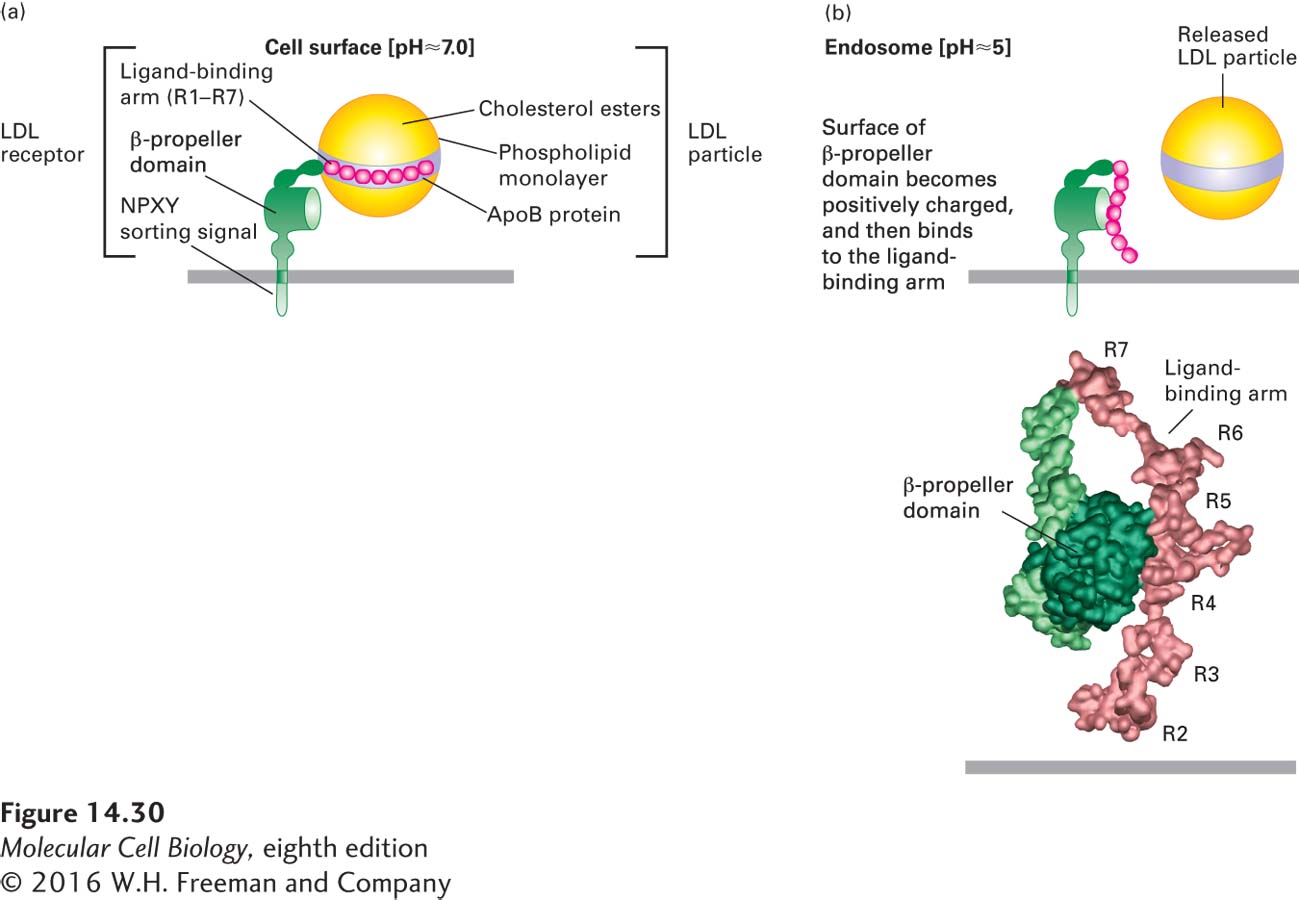
The mechanism by which the LDL receptor releases bound LDL particles is now understood in detail (Figure 14-30). At the endosomal pH of 5.0–5.5, histidine residues in a region known as the β-propeller domain of the receptor become protonated, forming a site that can bind with high affinity to the negatively charged cysteine-rich repeats in the LDL-binding domain. This intramolecular interaction sequesters the repeats in a conformation that cannot simultaneously bind to apoB-100, thus causing release of the bound LDL particle.
[Part (b) data from G. Rudenko et al., 2002, Science298:2353, PDB ID 1n7d.]
FIGURE 14-30Model for pH-dependent binding of LDL particles by the LDL receptor. Schematic depiction of an LDL receptor at the neutral pH found at the cell surface (a) and at the acidic pH found in the interior of the late endosome (b). (a) At the cell surface, apoB-100 on the surface of an LDL particle binds tightly to the receptor. Of the seven cysteine-rich repeats (R1–R7) in the ligand-binding arm, R4 and R5 appear to be most critical for LDL binding. (b, top) Within the endosome, histidine residues in the β-propeller domain of the LDL receptor become protonated. The positively charged propeller can bind with high affinity to the ligand-binding arm, which contains negatively charged residues, causing release of the LDL particle. (b, bottom) Experimental electron density and Cα backbone trace model of the extracellular region of the LDL receptor at pH 5.3 based on x-ray crystallographic analysis. In this conformation, extensive hydrophobic and ionic interactions occur between the β propeller and the R4 and R5 repeats.
[Part (b) data from G. Rudenko et al., 2002, Science298:2353, PDB ID 1n7d.]