Activated Phospholipase C Generates Two Key Second Messengers Derived from the Membrane Lipid Phosphatidylinositol 4,5-Bisphosphate
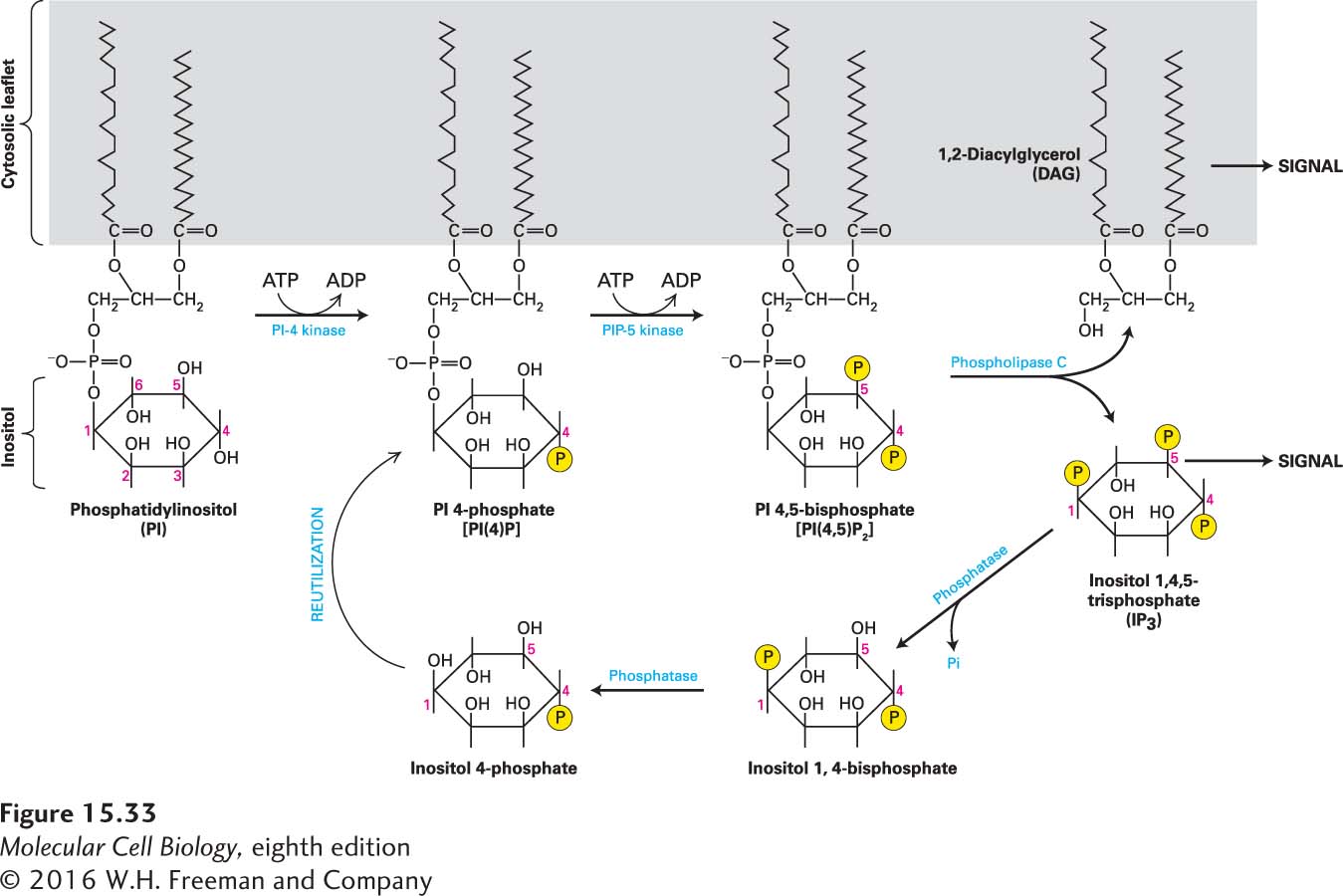
A number of important second messengers, used in several signal transduction pathways, are derived from the membrane lipid phosphatidylinositol (PI; Figure 15-33). The inositol group in this phospholipid, which always faces the cytosol, can be reversibly phosphorylated at one or more positions by the combined actions of various kinases and phosphatases discussed in Chapter 16. One derivative of PI, the lipid phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2], is made by stepwise addition of two phosphates to PI. PI(4,5)P2 is then cleaved by activated phospholipase C into two important second messengers: 1,2-diacylglycerol (DAG), a lipophilic molecule that remains associated with the membrane, and inositol 1,4,5-trisphosphate (IP3), which can freely diffuse in the cytosol (see Figure 15-33). We refer to downstream events involving these two second messengers collectively as the IP3/DAG pathway.
FIGURE 15-33Synthesis of second messengers DAG and IP3from phosphatidylinositol (PI). Each membrane-bound PI kinase places a phosphate (yellow circles) on a specific hydroxyl group on the inositol ring, producing the phosphorylated derivatives PI(4)P and PI(4,5)P2. Cleavage of PI(4,5)P2 by phospholipase C yields the two important second messengers DAG and IP3. Signaling is terminated when a phosphatase removes the 5-phosphate from IP3; a second phosphatase removes the 1-phosphate, and the inositol 4-phosphate is reused to synthesize PI 4-phosphate. See A. Toker and L. C. Cantley, 1997, Nature387:673, and C. L. Carpenter and L. C. Cantley, 1996, Curr. Opin. Cell Biol.8:153.
Page 710
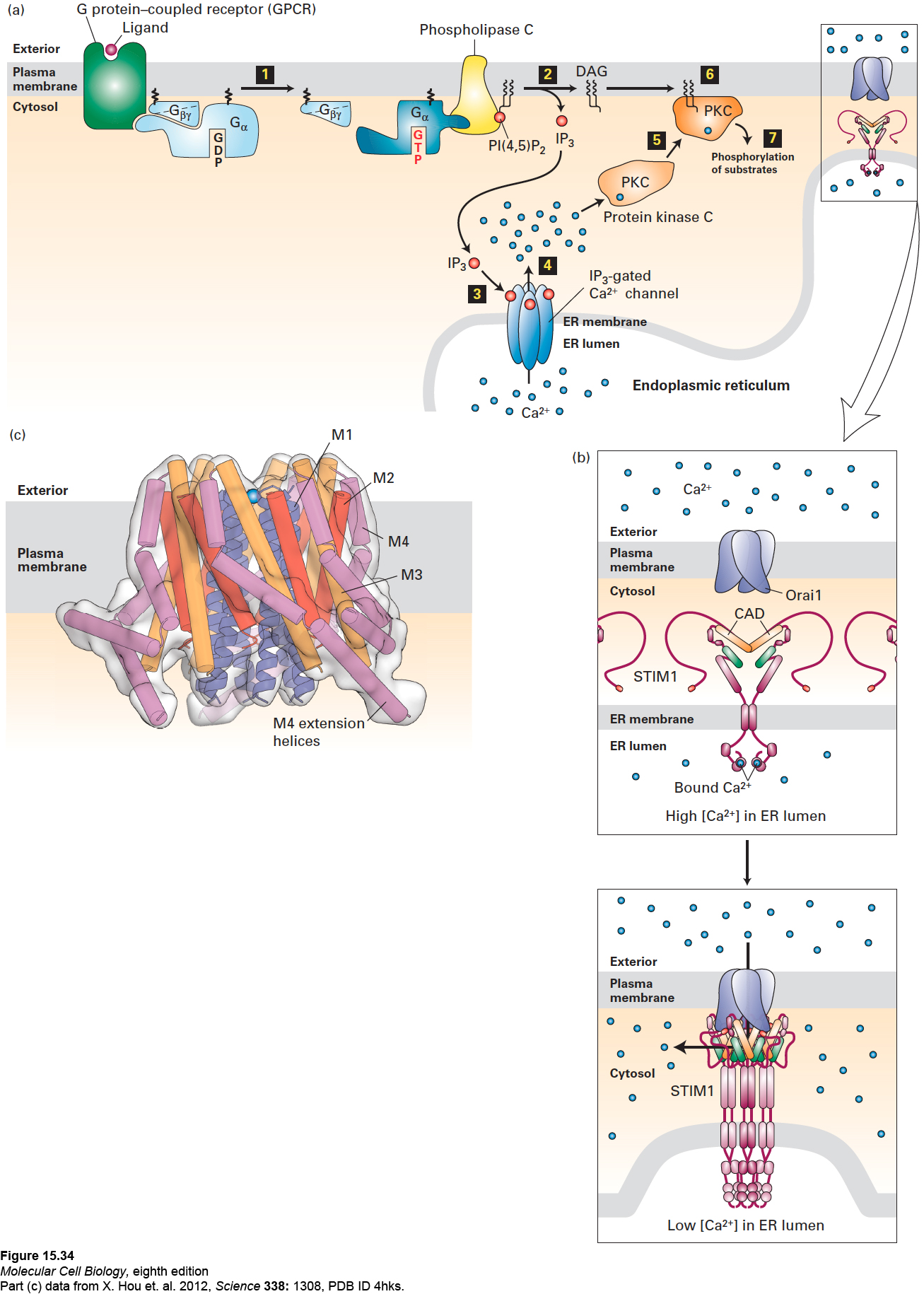
Phospholipase C is activated by G proteins containing either Gαo or Gαq subunits. In response to hormone activation of their associated GPCR, the Gαo or Gαq subunits bound to GTP separate from Gβγ and bind to and activate phospholipase C in the membrane (Figure 15-34a, step 1). In turn, activated phospholipase C cleaves PI(4,5)P2 into DAG and IP3 (see Figure 15-34a, step 2). The two second messengers trigger distinct, but sometimes interrelated, downstream effects.
Page 711
[Part (c) data from X. Hou et al. 2012, Science 338:1308, PDB ID 4hks.]
FIGURE 15-34The IP3/DAG pathway and the elevation of cytosolic Ca2+. (a) Opening of endoplasmic reticulum Ca2+ channels can be triggered by ligand binding to GPCRs that activate either the Gαo or Gαq subunit, leading to activation of phospholipase C (step 1). Cleavage of PI(4,5)P2 by phospholipase C yields IP3 and DAG (step 2). After diffusing through the cytosol, IP3 interacts with and opens IP3-gated Ca2+ channels in the membrane of the endoplasmic reticulum (step 3), causing release of stored Ca2+ ions into the cytosol (step 4). One of several cellular responses induced by a rise in cytosolic Ca2+ is recruitment of protein kinase C (PKC) to the plasma membrane (step 5), where it is activated by DAG (step 6). The activated membrane-associated kinase can phosphorylate various cellular enzymes and receptors, thereby altering their activity (step 7). (b) Opening of plasma-membrane Ca2+ channels. Top: In the resting cell, Ca2+ levels in the ER lumen are high, and Ca2+ ions (blue circles) bind to the luminal EF hand domains of the transmembrane STIM proteins. Bottom: As Ca2+ stores in the ER are depleted and Ca2+ ions dissociate from the EF hands, STIMs undergo oligomerization and relocalization to areas of the ER membrane near the plasma membrane. There the STIM CAD domains (orange) bind to and trigger the opening of the store-operated Ca2+ channels (Orai1) in the plasma membrane, allowing influx of extracellular Ca2+. (c) Drawing of the three-dimensional structure of Orai1 in the closed state. The Orai1 Ca2+ channel is composed of six identical subunits arranged around a central Ca2+ pore. Each subunit contains four transmembrane α helices—M1 (blue), M2 (red), M3 (orange), and M4 (violet)—and a helix following M4 that extends into the cytosol (termed the M4 extension helix, violet). The M1 helices are drawn as ribbons, the M2–M4 helices as cylinders. The pore is lined by the six M1 helices; in the closed state, a Ca2+ ion is bound at the extracellular entrance to the pore, but cannot enter it. Binding of the CADs leads to channel opening, most likely by widening of the pore by the outward movement of the M1 helices. The intracellular ends of the M1 helices are thought to interact with a portion of the STIM CAD, as are the M4 extensions, and it is hypothesized that the CADs bridge the cytosolic portions of the M1 helices and the M4/M4 extension helices. See J. W. Putney, 1999, P. Natl. Acad. Sci. USA 96:14669; Y. Zhou, 2010, P. Natl. Acad. Sci. USA107:4896; and M. Cahalan, 2010, Science330:43.
[Part (c) data from X. Hou et al. 2012, Science 338:1308, PDB ID 4hks.]
Page 712
Ca2+ Release from the ER Triggered by IP3 Ligand binding to G protein–coupled receptors that activate phospholipase C induces an elevation in cytosolic Ca2+ even when Ca2+ ions are absent from the surrounding extracellular fluid. In this case, Ca2+ is released into the cytosol from the ER lumen (where, as noted above, Ca2+ can accumulate to almost millimolar concentrations) through the operation of IP3-gated Ca2+channels in the ER membrane, as depicted in Figure 15-34a (steps 3 and 4). (This family of channel proteins is similar in structure to the family of voltage-sensitive Ca2+ channels that are called ryanodine receptors in muscle cell sarcoplasmic reticulum membranes; see page 805.) Each of these large IP3-gated channels is composed of four identical subunits, each of which contains an IP3-binding site in its N-terminal cytosolic domain. IP3 binding induces opening of the channel, allowing Ca2+ to flow down its concentration gradient from the ER lumen into the cytosol. When different phosphorylated inositols normally found in cells were added to preparations of ER vesicles, only IP3 caused release of Ca2+ ions from the vesicles. This simple experiment demonstrates the specificity of the IP3 effect.
The IP3-mediated rise in the cytosolic Ca2+ level is transient because Ca2+ pumps located in the plasma membrane and in the ER membrane actively transport Ca2+ from the cytosol to the cell exterior and back into the ER lumen, respectively. Furthermore, within a second of its generation, the phosphate linked to carbon 5 of IP3 is hydrolyzed, yielding inositol 1,4-bisphosphate (see Figure 15-33). This compound cannot bind to the IP3-gated Ca2+ channel and thus does not stimulate Ca2+ release from the ER.
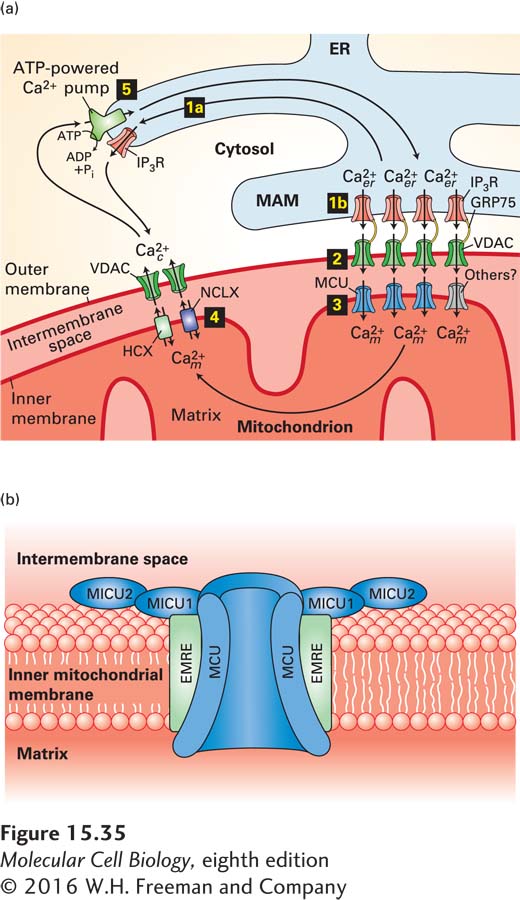
Ca2+ Transport from the ER to the Mitochondrial Matrix Triggered by IP3 In Chapter 12, we learned that direct contacts between specialized regions of the ER membrane, called mitochondria-associated membranes (MAMs), and the outer mitochondrial membrane affect mitochondrial structure, dynamics, and function. Regulated movement of Ca2+ from the ER lumen through the MAMs into the mitochondrial matrix is a major part of this regulation (Figure 15-35a). IP3-gated Ca2+ channels in the MAMs open in response to a rise in cytosolic IP3 (step 1a, Figure 15-35a). Voltage-dependent anion channels (VDACs) in the outer mitochondrial membrane adjacent to the MAMs, which are physically linked to these IP3-gated Ca2+ channels by the GRP75 protein, efficiently pass the Ca2+ released from the ER lumen into the intermembrane space (step 2). A mitochondrial calcium uniporter (MCU) in the inner mitochondrial membrane then transports Ca2+ into the mitochondrial matrix, where it increases ATP synthesis as well as enhancing mitochondrial activity in other ways.
FIGURE 15-35Movement of Ca2+between the cytosol, mitochondrion, and endoplasmic reticulum. (a) The ER is the main intracellular storage depot for Ca2+. Binding of IP3 to IP3-gated Ca2+ channels (IP3R) in the membrane of the endoplasmic reticulum releases Ca2+ into the cytosol (step 1a); binding also opens IP3-gated Ca2+ channels in the mitochondria-associated membranes (MAMs) of the ER (step 1b). Step 2: VDAC channels in the outer mitochondrial membrane adjacent to MAMs are physically linked to IP3Rs by the GRP75 protein; they efficiently pass the Ca2+ released from the MAMs into the intermembrane space. Step 3: The high concentration of Ca2+ in the intermembrane space induces the opening of MCUs or other Ca2+ channels in the inner membrane, resulting in the flow of Ca2+ into the mitochondrial matrix. Step 4: Over time, Ca2+ is released from the mitochondria by Ca2+/Na+ (NCLX) and Ca2+/H+ (HCX) antiporters in the inner membrane, then transferred into the cytosol through VDAC or other Ca2+ channels in the outer membrane. Finally, pumping of Ca2+ from the cytosol by ATP-powered Ca2+ pumps in the ER membrane (step 5) or plasma membrane restores the high ER Ca2+ and low cytosolic Ca2+ levels. (b) Model of the mitochondrial calcium uniporter complex. A multimer of MCU subunits forms the regulated Ca2+ pore. Additional subunits include the integral membrane protein EMRE and regulatory subunits MICU1 and MICU2. Ca2+ binding to the MICU subunits opens the MCU pore, resulting in the flow of the Ca2+ from the intermembrane space into the matrix. See M. Schäfer et al., 2014, Cell Tissue Res. 357:395 and K. Kamer and V. Mootha, 2015, Nat. Rev. Mol. Cell Biol. 16:545.
MCUs open only in the presence of a high Ca2+ concentration in the intermembrane space. Regulatory subunits of the MCU that face the intermembrane space (Figure 15-35b) have Ca2+-binding EF hands (see Chapter 3) with relatively low binding affinity for Ca2+; these subunits must bind calcium for the MCU to open. Individuals with mutations in the gene encoding one of these subunits have skeletal muscle defects and learning disabilities, symptoms that can accompany other mitochondrial disorders, attesting to the importance of these uniporters in mitochondrial metabolism.
Page 713
To avoid the buildup of excess, potentially toxic, intramitochondrial calcium, the mitochondrial matrix gradually releases Ca2+ into the cytosol. The calcium first moves across the inner mitochondrial membrane into the intermembrane space via Na+/Ca2+ and H+/Ca2+ antiporters, then crosses the outer mitochondrial membrane, most likely via VDACs (step 4). The calcium transport cycle is completed when cytosolic calcium enters the ER via an ATP-powered Ca2+ pump (step 5; see Figure 11-10) or is pumped out of the cell by plasma-membrane ATP-powered Ca2+ pumps.
The Store-Operated Plasma-Membrane Ca2+ Channel Continued opening of the IP3-gated Ca2+ channel in the ER membrane, coupled with operation of the plasma-membrane Ca2+ pump, would eventually deplete the intracellular stores of Ca2+, and a cell would soon be unable to increase the cytosolic Ca2+ level in response to hormone-induced IP3. Patch-clamping studies (see Figure 11-22) have revealed that a plasma-membrane Ca2+ channel called the store-operated channel opens in response to depletion of ER Ca2+ stores and admits extracellular Ca2+ into the cytosol. Studies in which each potential Ca2+ channel protein was knocked down one at a time with shRNAs (see Figure 6-42) established the identity of this channel protein as Orai1; the sequence and three-dimensional structure of Orai1 is unlike that of any other known ion-channel protein (see Figure 15-34c), which partly explains why it took so long to identify it as the store-operated channel.
The ER Ca2+-sensing protein is STIM, a transmembrane protein in the ER membrane (see Figure 15-34b). An EF hand, similar to that in calmodulin (see Figure 3-33), on the luminal side of the ER membrane binds Ca2+ when its level in the lumen is high. As stores of Ca2+ in the ER are depleted, the STIM proteins lose their bound Ca2+, oligomerize, and in an unknown manner, relocalize to areas of the ER membrane near the plasma membrane (see Figure 15-34b). There the CAD domains of the STIM proteins bind to and trigger the opening of Orai1, allowing the influx of extracellular Ca2+ (see Figure 15-34c). Combined overexpression of Orai and STIM in cultured cells leads to a marked increase in Ca2+ influx, establishing that these two proteins are the key components of the store-operated Ca2+ pathway.
[Data from A. Miyawaki et al., 1997, Nature388:882.]
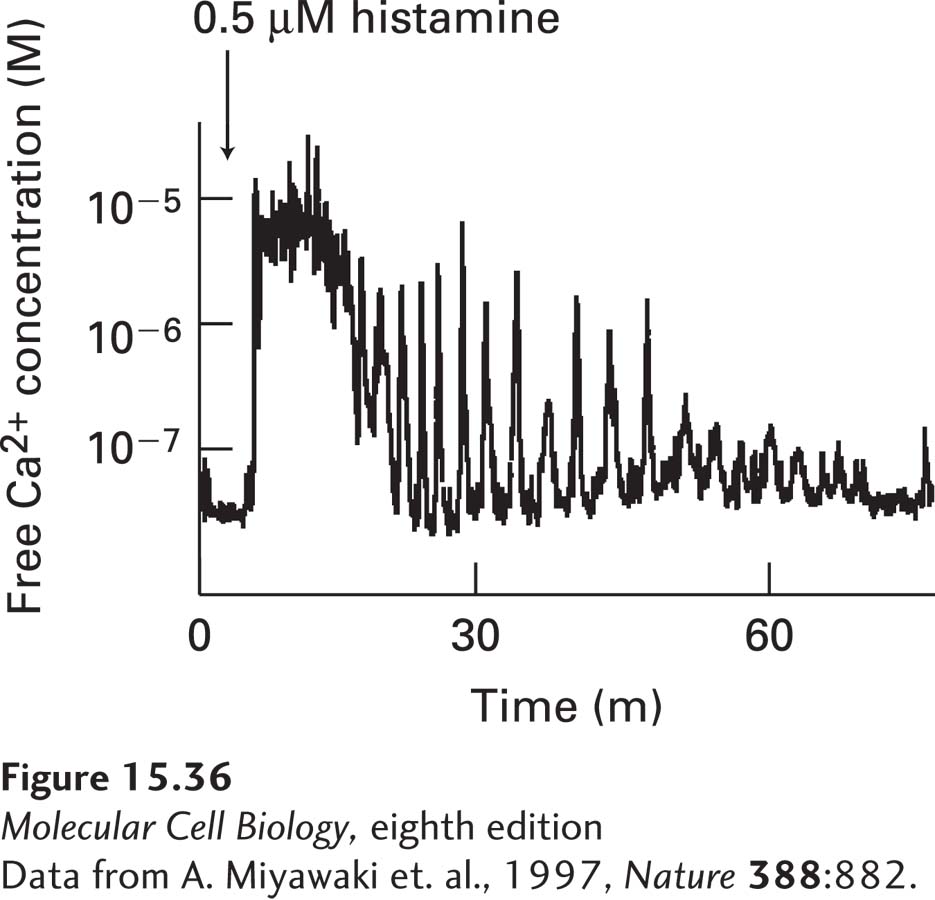
FIGURE 15-36Oscillations in the cytosolic Ca2+concentration following treatment of human HeLa cells with histamine. Like the LH receptors described in the text, the histamine GPCR activates the IP3-DAG signaling pathway. The feedback loops generating the spikes in the cytosolic Ca2+ concentration are detailed in the text.
[Data from A. Miyawaki et al., 1997, Nature388:882.]
Feedback Loops That Trigger Spikes in the Cytosolic Ca2+ Concentration Continuous activation of certain G protein–coupled receptors induces rapid, repeated spikes in the level of cytosolic Ca2+ (Figure 15-36). These oscillations in cytosolic Ca2+ levels are caused by a complex feedback interaction between the cytosolic Ca2+ concentration and the IP3-gated Ca2+-channels. The submicromolar level of cytosolic Ca2+ in the resting cell potentiates the opening of these channels via a rise in IP3, thus facilitating the rapid rise in cytosolic Ca2+ that follows hormone stimulation of cell-surface G protein–coupled receptors. However, the high cytosolic Ca2+ levels reached at the peak of the spike inhibit further IP3-induced release of Ca2+ from ER stores by decreasing the affinity of the Ca2+ channels for IP3. As a result, the IP3-gated Ca2+ channels close, and the cytosolic Ca2+ level drops rapidly as Ca2+ is pumped into the ER lumen or out of the cell. Thus cytosolic Ca2+ is a feedback inhibitor of the IP3-gated Ca2+ channels that, when open, trigger a rise in cytosolic Ca2+. As an example, this mechanism produces calcium ion oscillations occur in the pituitary gland cells that secrete luteinizing hormone (LH), which plays an important role in controlling ovulation and thus female fertility. LH secretion is induced by the binding of luteinizing hormone–releasing hormone (LHRH) to its G protein–coupled receptors on the surfaces of these cells; LHRH binding induces repeated Ca2+ spikes. Each Ca2+ spike induces exocytosis of a few LH-containing secretory vesicles, presumably those close to the plasma membrane.