GTP-Binding Proteins Are Frequently Used in Signal Transduction Pathways as On/Off Switches
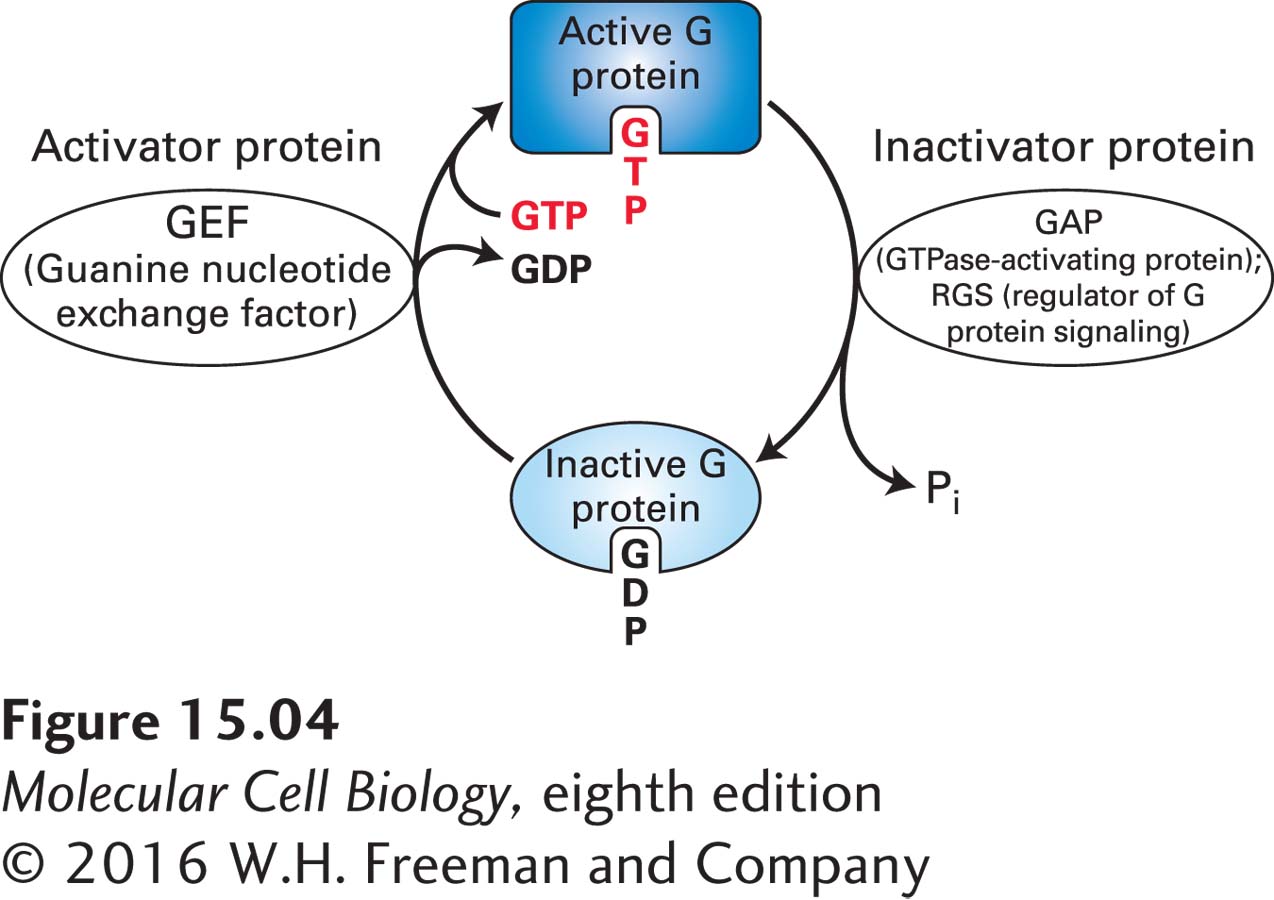
Many cellular processes utilize members of the GTPase superfamily of proteins, which are found in all prokaryotic and eukaryotic cells. All of these GTP-binding switch proteins exist in two forms (Figure 15-4): (1) an active (“on”) form with bound GTP (guanosine triphosphate), which modulates the activity of specific target proteins, and (2) an inactive (“off”) form with bound GDP (guanosine diphosphate), which cannot affect the activity of target proteins. Members of the GTPase superfamily switch between GTP-bound “on” and GDP-bound “off” forms. These proteins are evolutionarily ancient, as evidenced by their widespread functions in protein synthesis [examples include the roles of the eIF2 initiation factor (Figure 5-23) and the EF1α and EF2 proteins in protein elongation (Figure 5-24); the transport of proteins between the nucleus and the cytoplasm (Ran, Figure 13-36); the formation of transport vesicles (Sar proteins, Figure 14-8) and their fusion with target membranes (Rab proteins, Figure 14-10); and rearrangements of the actin cytoskeleton (Rho, Rac, and Cdc42 proteins, discussed in Chapter 17)].
FIGURE 15-4GTPase switch proteins cycle between active and inactive forms. The switch protein is active when it has bound GTP and inactive when it has bound GDP. Conversion of the active into the inactive form by hydrolysis of the bound GTP is accelerated by GAPs (GTPase-activating proteins), RGSs (regulators of G protein signaling), and other types of proteins. Reactivation is promoted by GEFs (guanine nucleotide exchange factors), which catalyze the dissociation of the bound GDP and its replacement by GTP.
Page 678
Here we focus on members of this superfamily that function in signal transduction pathways, in which conversion between the active GTP-bound and inactive GDP-bound states is tightly regulated. Conversion of the inactive to the active state is usually triggered by a signal (e.g., a hormone binding to a receptor) and is mediated by a guanine nucleotide exchange factor (GEF), which causes the release of GDP from the switch protein. Subsequent binding of GTP, favored by its high intracellular concentration relative to that of GDP, induces a conformational change to the active form. The principal conformational changes involve two highly conserved segments of the GTP-binding protein, termed switch I and switch II, that allow the protein to bind to and activate downstream signaling proteins (Figure 15-5). Conversion of the active form back to the inactive form is mediated by a GTPase, which is often part of the switch protein itself and which slowly hydrolyzes the bound GTP to GDP and Pi, thus altering the conformation of the switch I and switch II segments so that they are unable to bind to the target effector protein.
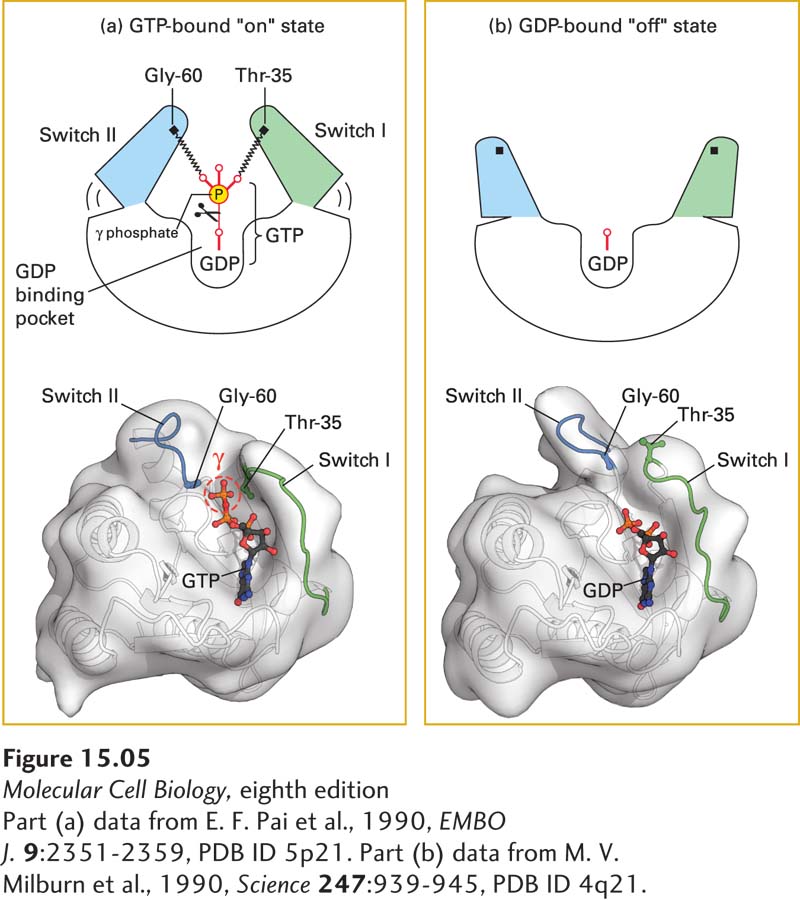
[Part (a) data from E. F. Pai et al., 1990, EMBO J. 9:2351-2359, PDB ID 5p21. Part (b) data from M. V. Milburn et al., 1990, Science247:939-945, PDB ID 4q21.]
FIGURE 15-5Switching mechanism of monomeric G proteins. The ability of a G protein to interact with other proteins and thus transduce a signal differs between the GTP-bound “on” state and GDP-bound “off” state. (a) In the active “on” state, two domains, termed switch I (green) and switch II (blue), are bound to the terminal γ phosphate of GTP through interactions with the backbone amide groups of conserved threonine and glycine residues. When bound to GTP in this way, the two switch domains are in a conformation such that they can bind to and thus activate specific downstream effector proteins. (b) Removal of the γ phosphate by GTPase-catalyzed hydrolysis causes switch I and switch II to relax into a different conformation, the inactive “off” state; in this state, they are unable to bind to effector proteins. The three-dimensional models shown here represent both conformations of Ras, a monomeric G protein. A similar spring-loaded mechanism switches the alpha subunit in heterotrimeric G proteins between the active and inactive conformations by movement of three, rather than two, switch segments.
[Part (a) data from E. F. Pai et al., 1990, EMBO J. 9:2351-2359, PDB ID 5p21. Part (b) data from M. V. Milburn et al., 1990, Science247:939-945, PDB ID 4q21.]
The rate of GTP hydrolysis regulates the length of time the switch protein remains in the active conformation and is able to signal its downstream target proteins: the slower the rate of GTP hydrolysis, the longer the protein remains in the active state. The rate of GTP hydrolysis is often modulated by other proteins. For instance, both GTPase-activating proteins (GAPs) and regulators of G protein signaling (RGSs) accelerate GTP hydrolysis (see Figure 15-4). Many regulators of G protein activity are themselves controlled by extracellular signals.
Two large classes of GTPase switch proteins are used in signaling. Heterotrimeric G proteins directly bind to and are activated by certain cell-surface receptors. As we will see in Section 15.3, G protein–coupled receptors function as guanine nucleotide exchange factors (GEFs), activating the heterotrimeric G protein to which they are coupled by triggering its release of GDP and binding of GTP. Monomeric (often called low-molecular-weight) G proteins, including Ras and various Ras-like proteins such as Ran and Sar, do not directly bind to receptors, but play crucial roles in many pathways that regulate cell division and cell motility, as is evidenced by the fact that mutations in genes encoding these G proteins frequently lead to cancer.