SH2 Domains in Action: JAK Kinases Activate STAT Transcription Factors
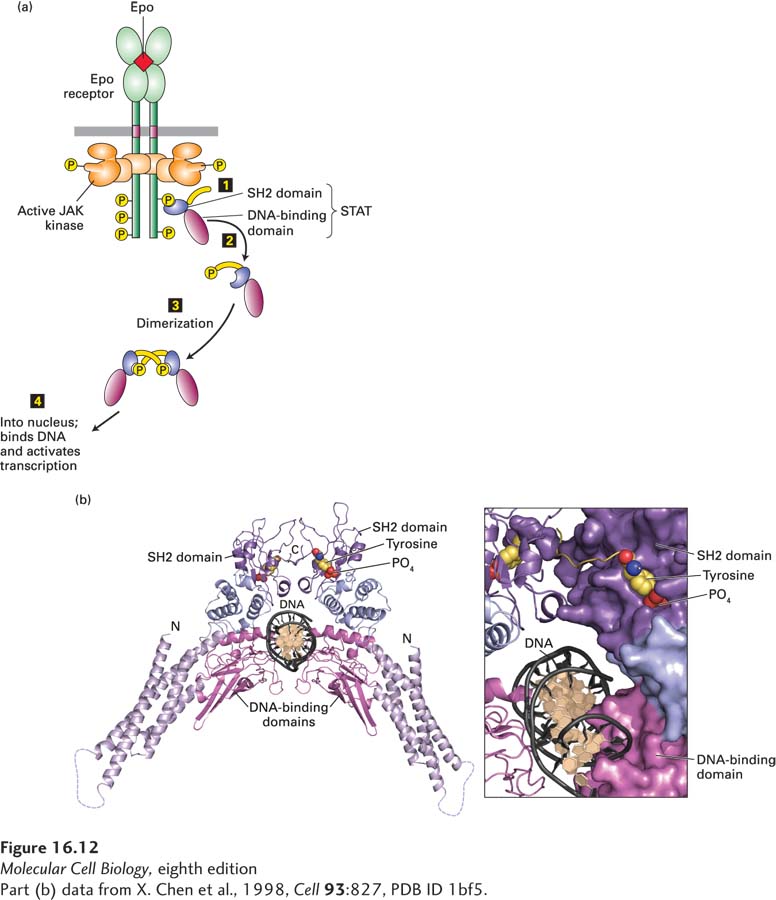
To illustrate how binding of SH2 domains to specific phosphotyrosine residues induces specific signaling pathways, let’s discuss the straightforward mechanism by which all JAK kinases, and some RTKs, directly activate members of the STAT family of transcription factors. All STAT proteins contain an N-terminal DNA-binding domain, an SH2 domain that binds to one or more specific phosphotyrosines in a cytokine receptor’s cytosolic domain, and a C-terminal domain with a critical tyrosine residue. Once a monomeric STAT is bound to a phosphotyrosine in the receptor via its SH2 domain, the C-terminal tyrosine is phosphorylated by the associated JAK kinase (Figure 16-12a). This arrangement ensures that in a particular cell, only those STAT proteins with an SH2 domain that can bind to a particular receptor will be activated, and only when that receptor is activated. For example, the erythropoietin receptor, as well as the receptors for GH, prolactin, G-CSF, and several more cytokines, activates STAT5, but not STATs 1, 2, 3, or 4; those STATs are activated by other cytokine receptors. A phosphorylated STAT dissociates spontaneously from the receptor, and two phosphorylated STAT proteins form a homodimer in which the SH2 domain on each binds to the phosphotyrosine in the other (Figure 16-12b). Because dimerization involves conformational changes that expose the nuclear-localization signal (NLS), the STAT dimers move into the nucleus, where they bind to specific enhancers or promoters (DNA regulatory sequences) controlling target genes (see Figure 16-12a) and thus alter gene expression.
[Part (b) data from X. Chen et al., 1998, Cell93:827, PDB ID 1bf5.]
FIGURE 16-12Activation and structure of STAT proteins. (a) Phosphorylation and dimerization of STAT proteins. Step 1: Following activation of a cytokine receptor (see Figure 16-10), the SH2 domain of an inactive monomeric STAT transcription factor binds to a phosphotyrosine in the receptor, bringing the STAT close to the active JAK associated with the receptor. The JAK then phosphorylates the C-terminal tyrosine in the STAT. Steps 2 and 3: Phosphorylated STATs spontaneously dissociate from the receptor and spontaneously dimerize. Because the STAT homodimer has two phosphotyrosine-SH2 domain interactions, whereas the receptor-STAT complex is stabilized by only one such interaction, phosphorylated STATs tend not to rebind to the receptor. Step 4: The STAT dimer moves into the nucleus, where it can bind to promoter sequences and activate transcription of target genes. (b) Ribbon diagram of the STAT1 dimer bound to DNA (black). The STAT1 dimer forms a C-shaped clamp around DNA that is stabilized by reciprocal and highly specific interactions between the SH2 domain (purple) of one monomer and the phosphorylated tyrosine residue (yellow with red oxygens) on the C-terminal segment of the other. The phosphotyrosine-binding site of the SH2 domain in each monomer is coupled structurally to the DNA-binding domain (magenta), suggesting a potential role for the SH2-phosphotyrosine interaction in the stabilization of DNA interacting elements.
[Part (b) data from X. Chen et al., 1998, Cell93:827, PDB ID 1bf5.]
Because different cell types have unique complements of transcription factors and unique epigenetic modifications of their chromatin, the genes that are available to be activated by any STAT are different in different cell types. For example, in mammary gland cells, STAT5, the same STAT that is activated by EpoR in erythroid progenitor cells, becomes activated following prolactin binding to the prolactin receptor and induces transcription of genes encoding milk proteins. In contrast, when STAT5 becomes activated in erythroid progenitor cells following binding of Epo to EpoR, it induces expression of the protein Bcl-xL. Bcl-xL prevents the programmed cell death, or apoptosis, of these progenitors (see Chapter 21), allowing them to proliferate and differentiate into red blood cells. More generally, it is thought that in each type of cell, activated STAT proteins, like the Smads discussed earlier, bind only to DNA sites in open chromatin and mainly to sites that have master transcription factors or other cell-specific gene regulatory proteins bound at adjacent sites. This combinatorial diversity allows a relatively limited set of receptors, JAK kinases, and STAT proteins to control a vast array of cellular activities.