Concentration Gradients of Wnt Protein Are Essential for Many Steps in Development
Wnts are secreted proteins; however, in part because of the hydrophobic lipid that is covalently linked to these proteins, they diffuse only a short distance from a signaling cell and generally have localized effects. As Wnt diffuses farther and farther away from secreting cells, its concentration decreases. Different Wnt concentrations induce different fates in target cells: cells that receive a large amount of Wnt turn on certain genes and form certain structures; cells that receive a smaller amount turn on different genes and so form different structures. Signals that induce different cell fates depending on their concentration at their target cells are referred to as morphogens.
Perhaps the most striking example of Wnt as a morphogen occurs during regeneration of the planarian Schmidtea mediterranea (see Figure 1-22e). If the planarian’s head is cut off, a new one regenerates within 14 days; a normal, albeit smaller, worm is regenerated. Similarly, after removal of the tail, a new one regenerates. Most strikingly, a small body piece from the middle of the animal regenerates a normal head from the anterior (head-
Page 754
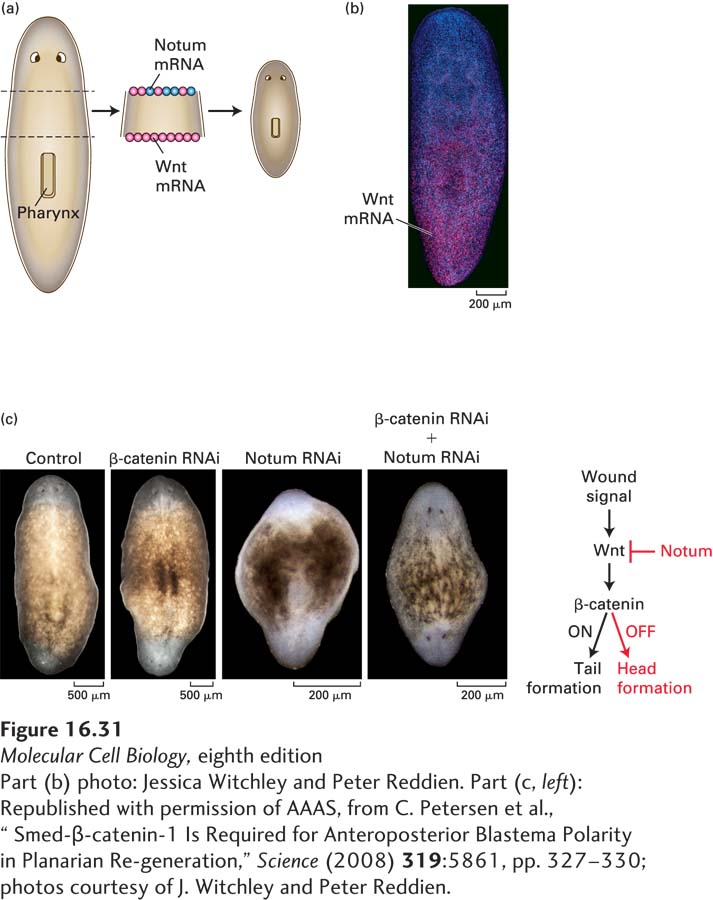
Figure 16-31c shows some elegant experiments establishing that signaling at the posterior wound by Wnt causes induction of a tail and that it is the absence of Wnt signaling at the anterior wound, rather than the presence of Notum, that induces head formation:
In the absence of all Wnt signaling, caused by treatment of the excised body fragment with an inhibitory RNA specific for β-catenin, a two-
headed planarian is regenerated; in this case, Wnt signaling is absent at both the anterior and posterior wounds, and thus heads are induced at both wound sites. Conversely, in the absence of Notum signaling, caused by treatment with an inhibitory RNA specific for Notum, a two-
tailed planarian is regenerated; in this case, Wnt is present and active at both the anterior and posterior wounds, and thus two tails are formed. In the absence of both Wnt and Notum signaling, caused by treatment with inhibitory RNAs for both β-catenin and Notum, the phenotype is similar to that caused by the loss of Wnt alone: a two-
headed planarian is regenerated. Wnt signaling is absent at both the anterior and posterior wounds, inducing head regeneration at both sites, and the absence of Notum is irrelevant in the absence of Wnt. Thus it is the absence of Wnt/β-catenin signaling, not the expression of Notum, that causes a tail to be formed.
These experiments support the role of Wnt concentration gradients in planarian regeneration. In Chapter 21, we will learn that mature planaria contain multipotent stem cells termed neoblasts that can differentiate into any body cell type; clearly gradients of Wnt protein play a major role in instructing neoblasts to differentiate into the multiple cell types that constitute the planarian head or tail.