Regulated Intramembrane Proteolysis of SREBPs Releases a Transcription Factor That Acts to Maintain Phospholipid and Cholesterol Levels
Although this chapter is focused on signaling pathways initiated by extracellular molecules, signaling pathways that sense the levels of internal molecules and respond accordingly sometimes share principles of molecular regulation and even mechanisms with pathways initiated from outside the cell. One such case is the control of cellular membrane lipids. A cell would soon face a crisis if it did not have enough phospholipids to make adequate amounts of membranes or had so much cholesterol that large crystals formed and damaged cellular structures (see Chapter 7). Cells sense the relative amounts of cholesterol and phospholipids in their membranes; they respond by adjusting the rates of cholesterol biosynthesis and import so that the cholesterol:phospholipid ratio is kept within a narrow desirable range. Regulated intramembrane proteolysis plays an important role in this cellular response to altered cholesterol levels.
Page 764
As we learned in Chapter 14, low-density lipoprotein (LDL) is rich in cholesterol and functions in transporting this lipid through the aqueous circulatory system (see Figure 14-27). Both enzymes in the cholesterol biosynthetic pathway (see Figure 7-26) and cellular levels of the LDL receptors that mediate cellular uptake of LDL are down-regulated when cellular cholesterol levels are adequate. Because LDL is imported into cells via receptor-mediated endocytosis (see Figure 14-29), a decrease in the number of LDL receptors leads to reduced cellular import of cholesterol.
Both cholesterol biosynthesis and cholesterol import are regulated at the level of gene transcription. For example, when growing cultured cells that need new membrane for sustained division are incubated with an external source of cholesterol, such as LDL, the level and the activity of HMG-CoA reductase, the rate-controlling enzyme in cholesterol biosynthesis, are suppressed. In contrast, the activity of acyl:cholesterol acyl transferase (ACAT), the enzyme that converts cholesterol into the esterified storage form, is increased. Thus energy is not wasted making unnecessary additional cholesterol, and cholesterol homeostasis is achieved.
Genes whose expression is controlled by the level of sterols such as cholesterol often contain one or more 10-base-pair sterol regulatory elements (SREs), or SRE half-sites, in their promoters. (These SREs differ from the serum response elements that control many early response genes, discussed in Section 16.4.) The interaction of cholesterol-dependent transcription factors called SRE-binding proteins (SREBPs) with these response elements modulates the expression of the target genes. How do cells sense how much cholesterol they have, and how is this “signal” used to control the level of SREBPs in the nucleus and thus gene expression? The SREBP-mediated pathway begins in the membranes of the endoplasmic reticulum (ER) and includes at least two other proteins besides SREBP.
Page 765
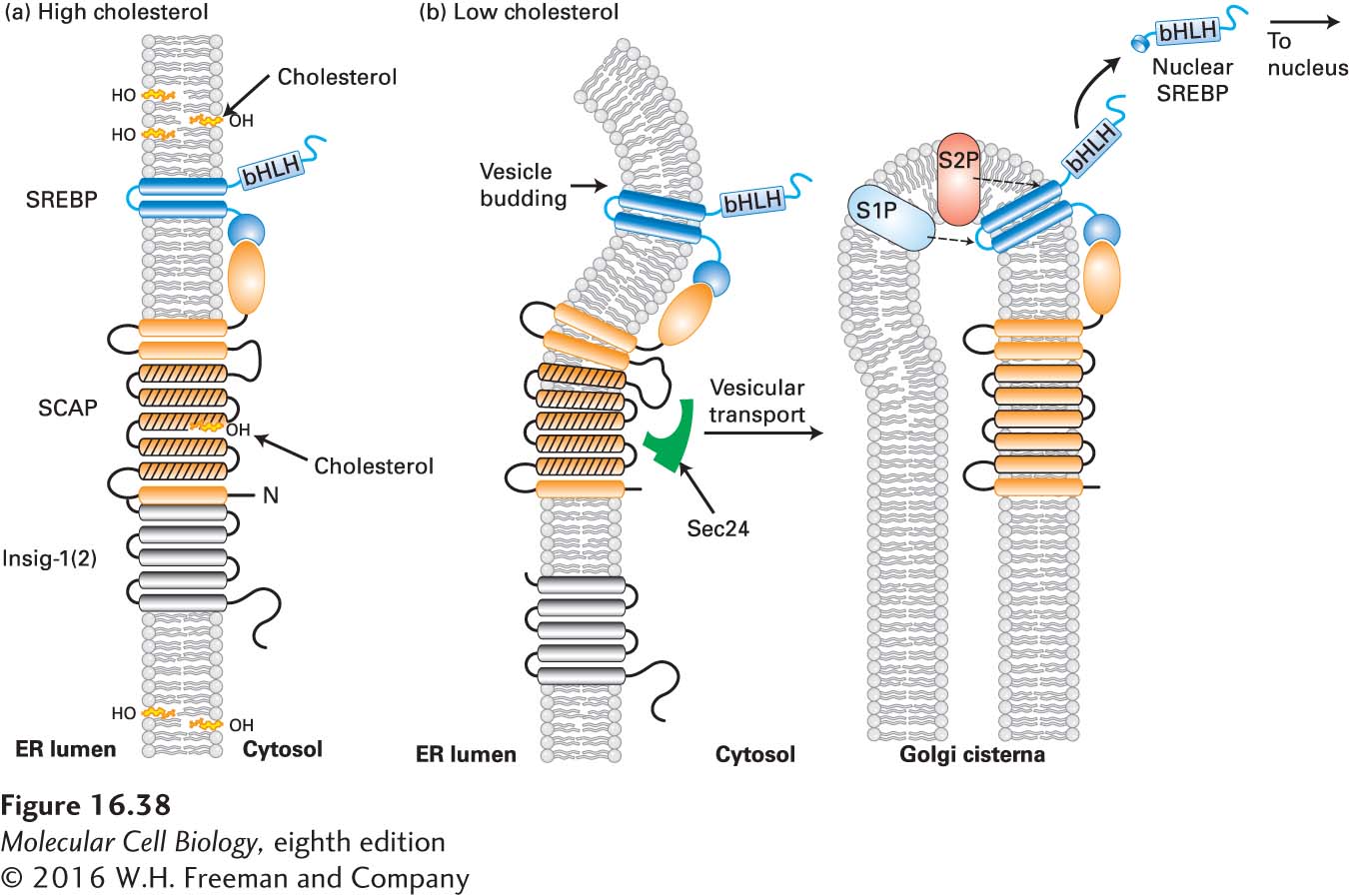
When cells have adequate concentrations of cholesterol, SREBP is found in the ER membrane complexed with SCAP (SREBP cleavage-activating protein), insig-1 (or its close homolog insig-2), and perhaps other proteins (Figure 16-38a). SREBP has three distinct domains: an N-terminal cytosolic domain, containing a basic helix-loop-helix (bHLH) DNA-binding motif (see Figure 9-30d), that functions as a transcription factor when cleaved from the rest of SREBP; a central membrane-anchoring domain containing two transmembrane α helices; and a C-terminal cytosolic regulatory domain. SCAP has eight transmembrane α helices and a large C-terminal cytosolic domain that interacts with the regulatory domain of SREBP. Five of the transmembrane α helices in SCAP form a sterol-sensing domain similar to that in HMG-CoA reductase (Figure 16-38a; see Section 7.3). When the sterol-sensing domain in SCAP is bound to cholesterol, the protein also binds to insig-1(2). When insig-1(2) is tightly bound to the SCAP-cholesterol complex, it blocks the binding of SCAP to the Sec24 coat protein subunit of COPII vesicles, thereby preventing incorporation of the SCAP-SREBP complex into ER-to-Golgi transport vesicles (see Chapter 14). This occurs when cholesterol concentrations in the ER membrane exceed 5 percent of total ER membrane lipids. Thus the cholesterol-dependent binding of insig-1(2) to the SCAP-cholesterol-SREBP complex traps that complex in the ER.
FIGURE 16-38Cholesterol-sensitive control of SREBP activation. The cellular pool of cholesterol is monitored by the combined action of insig-1(2) and SCAP, both transmembrane proteins located in the ER membrane. Membrane-spanning helices 2–6 of SCAP (orange with black lines) form a sterol-sensing domain, and a C-terminal segment binds to SREBP. (a) When cholesterol levels are high enough that ER cholesterol exceeds 5 percent of total ER lipids, cholesterol binds to the sterol-sensing domain in SCAP, triggering a conformational change that enables the N-terminal SCAP domain to bind to insig-1(2), anchoring the SCAP–SREBP complex in the ER membrane. (b) At low cholesterol levels, cholesterol dissociates from the SCAP sterol-sensing domain, triggering a reverse conformational change that dissociates SCAP from insig-1(2) and enables SCAP to bind to Sec24, a subunit of the COPII complex (see Figure 14-8). This binding initiates movement of the SCAP-SREBP complex to the Golgi complex by vesicular transport. In the Golgi, the sequential cleavage of SREBP by the site 1 and site 2 proteases (S1P, S2P) releases the N-terminal bHLH domain of SREBP, which translocates to the nucleus, and SCAP, which recycles to the ER. In the nucleus, the released SREBP domain, called nuclear SREBP (nSREBP), controls the transcription of genes containing sterol regulatory elements (SREs) in their promoters. See A. Radhakrishnan, 2008, Cell Metab.8:451, and M. Brown and J. Goldstein, 2009, J. Lipid Res.50:S15.
Cholesterol bound to SCAP is released when cellular cholesterol levels drop to less than 5 percent of ER lipids, a value that reflects total cellular cholesterol levels. Consequently, insig-1(2) no longer binds to the cholesterol-free SCAP, and the SCAP–SREBP complex moves from the ER to the Golgi complex via COPII vesicles (Figure 16-38b). SREBP is cleaved sequentially at two sites by two proteases in the Golgi membrane, S1P and S2P; the second cleavage represents an additional example of regulated intramembrane proteolysis. This second cleavage at site 2 releases the N-terminal bHLH-containing domain into the cytosol. This fragment, called nSREBP (nuclear SREBP), is rapidly translocated into the nucleus. There it activates transcription of genes containing sterol regulatory elements (SREs) in their promoters, such as those encoding the LDL receptor and HMG-CoA reductase. Thus a reduction in cellular cholesterol, by activating the insig-1(2)/SCAP/SREBP pathway, triggers expression of genes encoding proteins that both import cholesterol into the cell (the LDL receptor) and synthesize cholesterol from small precursor molecules (HMG-CoA reductase).
Page 766
After cleavage of SREBP in the Golgi, SCAP apparently recycles back to the ER, where it can interact with insig-1(2) and another intact SREBP molecule. High-level transcription of SRE-controlled genes requires the ongoing generation of new nSREBP because it is degraded fairly rapidly by the ubiquitin-mediated proteasomal pathway (see Chapter 3). The rapid generation and degradation of nSREBP help cells respond quickly to changes in levels of intracellular cholesterol.
Under some circumstances (e.g., during cell growth), cells need an increased supply of all the essential membrane lipids and their fatty acid precursors (which requires coordinate regulation). To make steroid hormones, cells sometimes need greater amounts of some lipids (such as cholesterol) than others, such as phospholipids (which requires differential regulation). How is such differential production achieved? Mammals express three known isoforms of SREBP: SREBP-1a and SREBP-1c, which are generated from alternatively spliced RNAs produced from the same gene, and SREBP-2, which is encoded by a different gene. Together, these intramembrane cleavage–activated transcription factors control expression of proteins that regulate availability not only of cholesterol, but also of fatty acids and the triglycerides and phospholipids made from fatty acids. In mammalian cells, SREBP-1a and SREBP-1c exert a greater influence on fatty acid metabolism than on cholesterol metabolism, whereas the reverse is the case for SREBP-2.