Myosin Heads Take Discrete Steps Along Actin Filaments
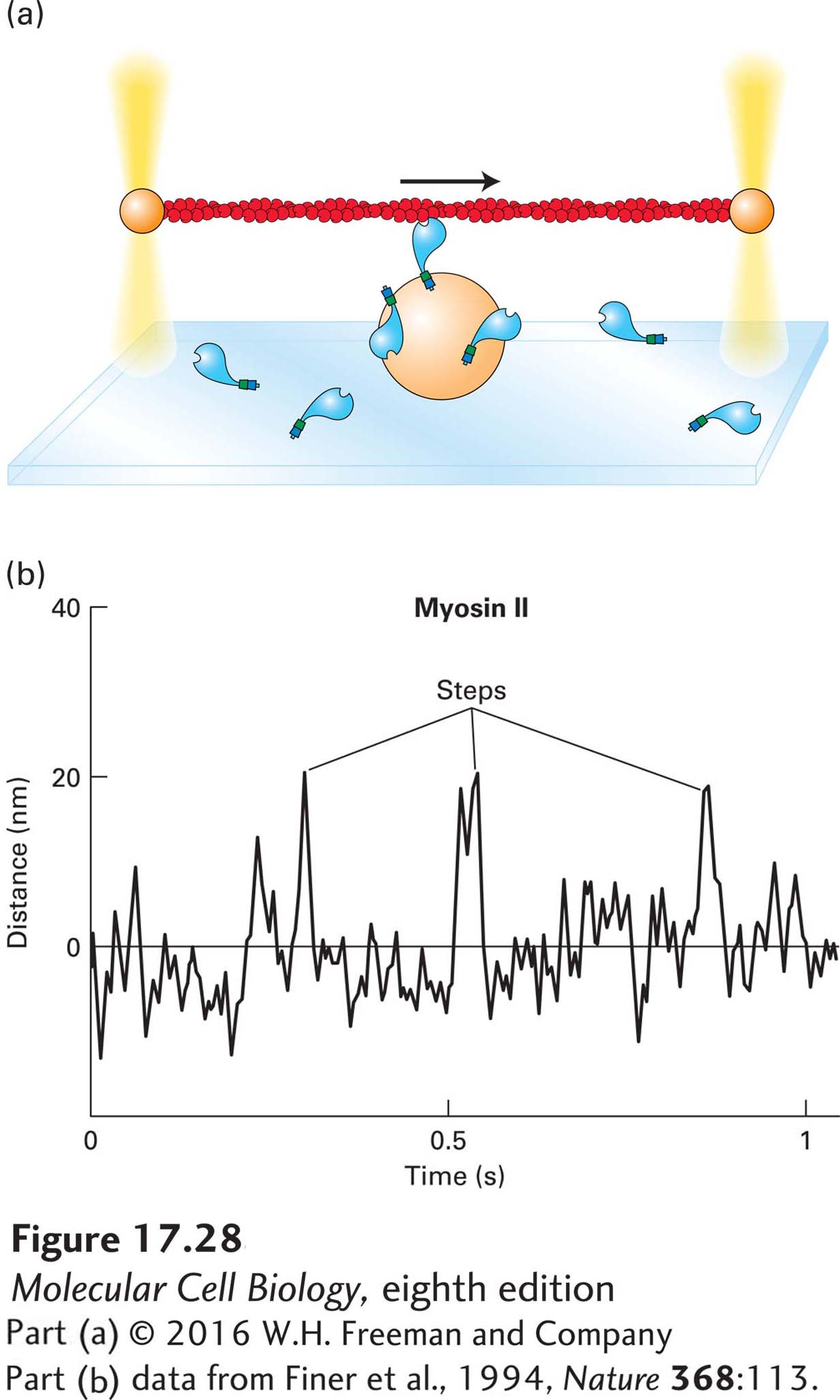
The most critical feature of myosin is its ability to generate a force that powers movements. Researchers have used optical traps to measure the forces generated by single myosin molecules (Figure 17-28a). In this approach, myosin is immobilized on beads at a low density. An actin filament, held between two optical traps, is lowered toward the bead until it contacts a myosin molecule on the bead. When ATP is added, the myosin pulls on the actin filament. Using a mechanical feedback mechanism controlled by a computer, one can measure the distance pulled and the forces and duration of the movement. The results of optical trap studies show that myosin II takes discrete steps, which average out to about 8 nm (Figure 17-28b), and generates 3–5 piconewtons (pN) of force, approximately the same force as that exerted by gravity on a single bacterium. We can also see that myosin II does not interact with the actin filament continuously, but rather binds, moves, and releases it (see Figure 17-28b). In fact, myosin II spends on average only about 10 percent of each ATPase cycle in contact with F-actin—it is said to have a duty ratio of 10 percent. This observation will be important later when we consider that in contracting muscle, hundreds of myosin heads pull on actin filaments, so that at any one time, 10 percent of the heads are engaged to provide a smooth contraction.
[Part (b) data from Finer et al., 1994, Nature368:113.]
EXPERIMENTAL FIGURE 17-28Measuring myosin step size and force with actin held by optical traps. (a) Optical trap techniques can be used to determine the step size and force generated by a single myosin molecule. In an optical trap, the beam of an infrared laser is focused by a light microscope on a latex bead (or any other object that does not absorb infrared light) to capture and hold the bead in the center of the beam. The strength of the force holding the bead is adjusted by increasing or decreasing the intensity of the laser beam. In this experiment, an actin filament is held between two optical traps. The actin filament is then lowered onto a third bead coated with a dilute concentration of myosin molecules. If the actin filament encounters a myosin molecule in the presence of ATP, the myosin will pull on the actin filament, which allows the investigators to measure both the force generated and the step size the myosin takes. (b) Using an optical trap setup, investigators have analyzed the behavior of myosin II. As shown by the peaks in the trace, myosin II takes erratic small steps (5–15 nm), which means that it binds the actin filament, moves, and then lets go. It is therefore a nonprocessive motor.
[Part (b) data from Finer et al., 1994, Nature368:113.]
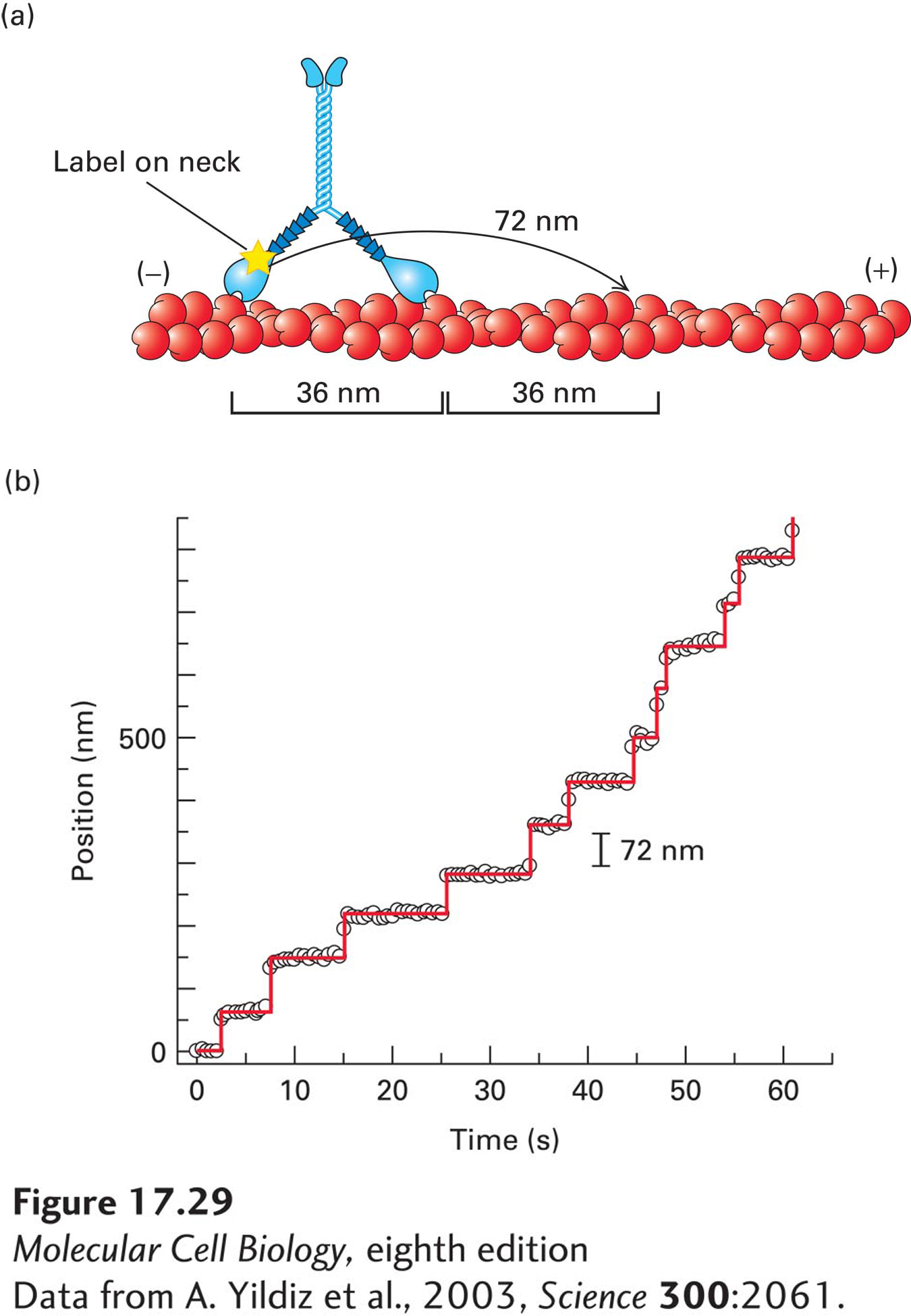
Now let us examine how myosin V moves. Scientists have managed to attach a fluorescent probe to just one of the two neck regions of a myosin V molecule and watch the fluorescent image as the molecule moves along an actin filament (Figure 17-29a). The labeled head takes many 72-nm steps without releasing from the actin—it is said to move processively (Figure 17-29b). This step size is twice the 36-nm length between helical repeats in the actin filament (see Figures 17-5b and 17-29a). So each 36-nm helical repeat site on the filament presents binding sites as each head alternately takes 72-nm steps—like someone walking on stepping stones across a river and placing each foot on every other stone. Notice that although the step size for each individual head is 72 nm, a cargo attached to the myosin V tail region will move 36 nm per step as the head moves from behind the cargo to in front of it. Thus the step size of the motor overall is said to be 36 nm, as indicated in Figure 17-29. Myosin V has presumably evolved to have a long neck domain—the lever arm—to take large steps to match the size of the helical repeat of the filament. Moreover, its ATPase cycle has been modified to have a much higher duty ratio (>70 percent) by slowing the rate of ADP release; thus the head remains in contact with the actin filament for a much larger percentage of the cycle. Since a single myosin V molecule has two heads, a duty ratio greater than 50 percent ensures that it maintains contact at all times as it moves down an actin filament, so that it does not fall off. These are exactly the properties one would expect for a motor designed to transport cargo along an actin filament.
Page 803
[Data from A. Yildiz et al., 2003, Science300:2061.]
EXPERIMENTAL FIGURE 17-29Myosin V has a step size of 36 nm, with each head stepping hand-over-hand in 72-nm steps. (a) Researchers have managed to label the neck of just one head of myosin V and follow its movement down an actin filament with nanometer accuracy (see Section 4.2 for how this can be achieved). (b) Trace of the label on one myosin V molecule as it walks down an actin filament. The labeled myosin head takes successive 72-nm steps. When the label was attached to the tail, the myosin V motor as a whole was found to take 36-nm steps (not shown). Thus myosin V heads step hand-over-hand down an actin filament in 72-nm steps for each head, but with the motor as a whole moving 36 nm per step. As can be seen from the trace, myosin V takes many successive steps along a filament, so it is said to be processive. As shown in panel (a), the step size corresponds to equivalent sites on the helical structure of the actin filament.
[Data from A. Yildiz et al., 2003, Science300:2061.]