Myosin Thick Filaments and Actin Thin Filaments in Skeletal Muscle Slide Past Each Other During Contraction
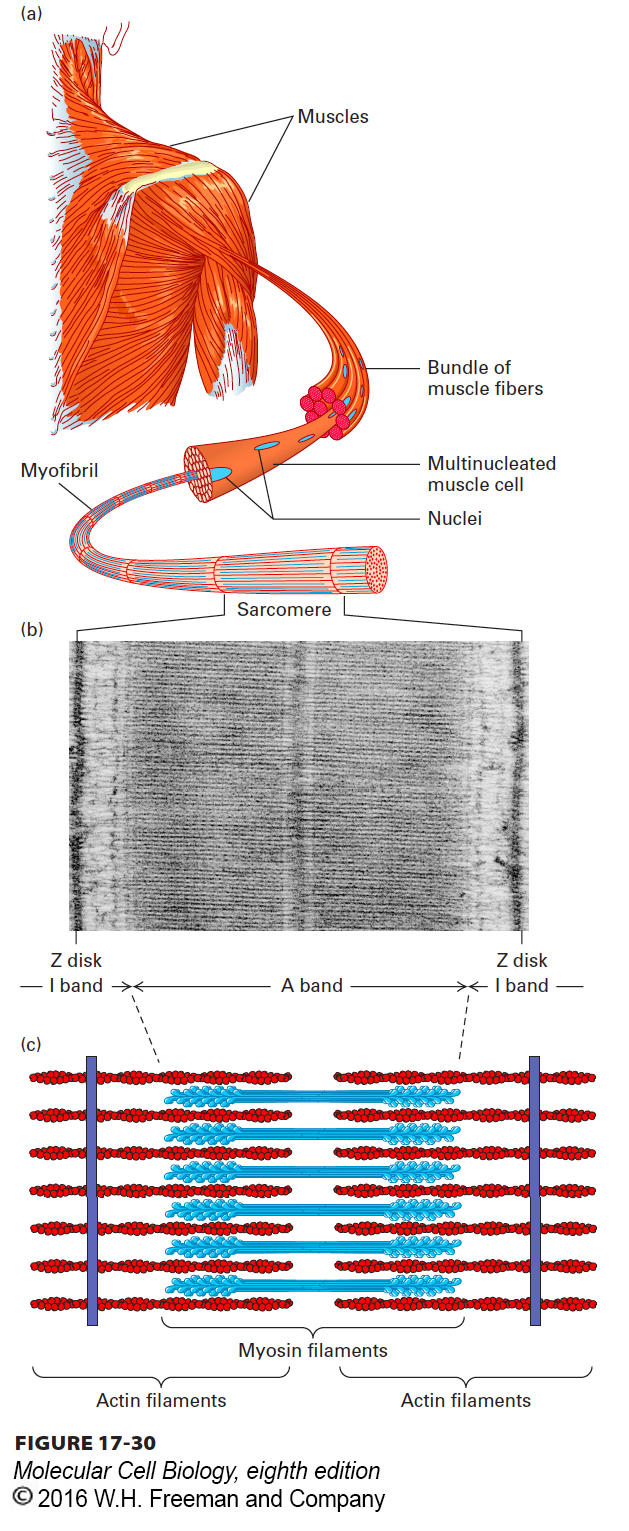
Muscle cells have evolved to carry out one highly specialized function: contraction. Muscle contractions must occur quickly and repetitively, and they must occur over long distances and with enough force to move large loads. A typical skeletal muscle cell is cylindrical, large (1–40 mm in length and 10–50 µm in width), and multinucleated (containing as many as 100 nuclei) (Figure 17-30a). Within each muscle cell are many myofibrils consisting of a regular repeating array of a specialized structure called a sarcomere (Figure 17-30b). A sarcomere, which is about 2 µm long in resting muscle, shortens by about 70 percent of its length during contraction. Electron microscopy and biochemical analysis have shown that each sarcomere contains two major types of filaments: thick filaments, composed of myosin II, and thin filaments, containing actin and associated proteins (Figure 17-30c).
FIGURE 17-30Structure of the skeletal muscle sarcomere. (a) Skeletal muscles consist of muscle fibers made of bundles of multinucleated cells. Each cell contains a bundle of myofibrils, which consist of thousands of repeating contractile structures called sarcomeres. (b) Electron micrograph of mouse skeletal muscle in longitudinal section, showing one sarcomere. On either side of the Z disks are the lightly stained I bands, composed entirely of actin thin filaments. These thin filaments extend from both sides of the Z disk to interdigitate with the dark-stained myosin thick filaments in the A band. (c) Diagram of the arrangement of myosin and actin filaments in a sarcomere.
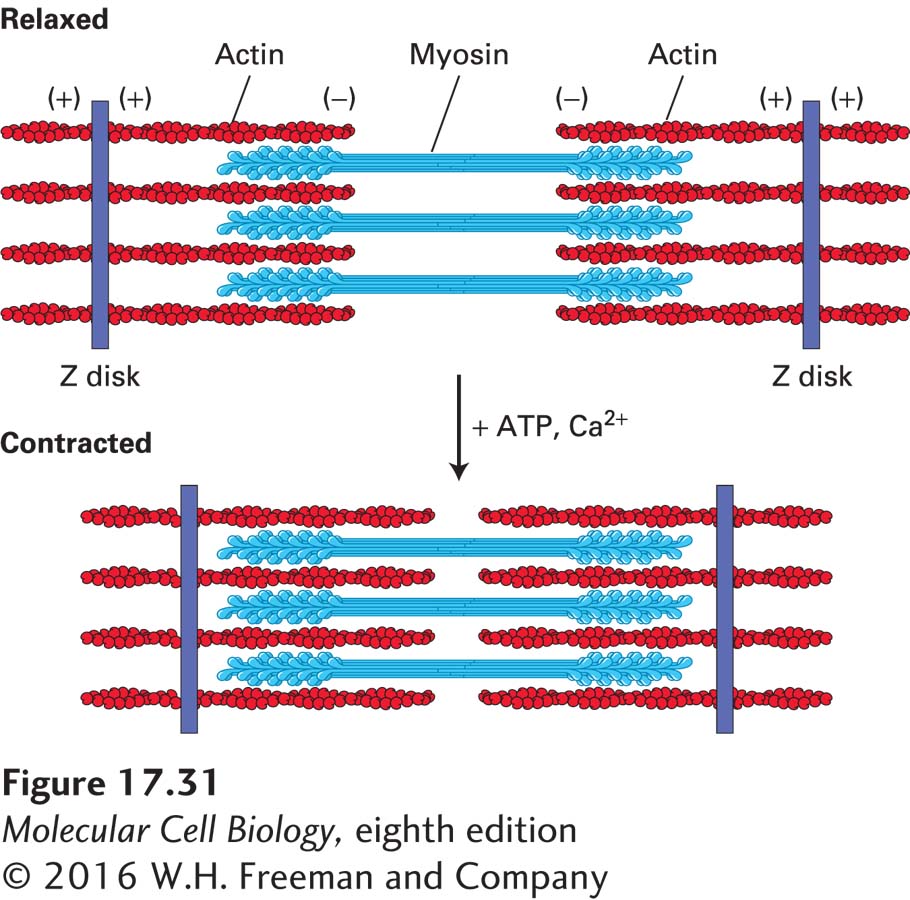
The thick filaments are myosin II bipolar filaments, in which the heads on each half of the filament have opposite orientations (see Figure 17-22a). The thin actin filaments are assembled with their (+) ends embedded in a densely staining structure known as the Z disk, so that the two sets of actin filaments in a sarcomere have opposite orientations (Figure 17-31). To understand how a muscle contracts, consider the interactions between one myosin head (among the hundreds in a thick filament) and one thin (actin) filament, as diagrammed in Figure 17-26. During these cyclical interactions, also called the cross-bridge cycle, the hydrolysis of ATP is coupled to the movement of a myosin head toward the Z disk, which corresponds to the (+) end of the actin thin filament. Because the thick filament is bipolar, the action of the myosin heads at opposite ends of the thick filament draws the thin filaments toward the center of the thick filament and therefore toward the center of the sarcomere (see Figure 17-31). This movement shortens the sarcomere until the ends of the thick filaments abut the Z disk. Contraction of an intact muscle results from the activity of hundreds of myosin heads on a single thick filament, amplified by the hundreds of thick and thin filaments in a sarcomere and thousands of sarcomeres in a muscle fiber. We can now see why myosin II is both nonprocessive and needs to have a low duty ratio: each head pulls a short distance on the actin filament and then lets go to allow other heads to pull, and so many heads working together allow the smooth contraction of the sarcomere. The first experimental basis for the sliding-filament model of muscle contraction is highlighted in Classic Experiment 17-1.
FIGURE 17-31The sliding-filament model of contraction in skeletal muscle. The arrangement of thick myosin and thin actin filaments in the relaxed state is shown in the top diagram. In the presence of ATP and Ca2+, the myosin heads extending from the thick filaments walk toward the (+) ends of the thin filaments. Because the thin filaments are anchored at the Z disks (purple), movement of myosin pulls the actin filaments toward the center of the sarcomere, shortening its length in the contracted state, as shown in the bottom diagram.
The human heart is an amazing contractile organ—it contracts without interruption about 3 million times a year, or a fifth of a billion times in a lifetime. The muscle cells of the heart contain contractile machinery very similar to that of skeletal muscle, except that they are mono- and bi-nucleated cells. In each cell, the end sarcomeres insert into structures at the plasma membrane called intercalated disks, which link the cells into a contractile chain. Heart muscle cells are generated only early in life, so they cannot be replaced in response to damage, such as occurs during a heart attack. Many different mutations in proteins of the heart contractile machinery give rise to hypertrophic cardiomyopathies—thickening of the heart wall muscle, which compromises its function. For example, many mutations have been documented in the cardiac myosin heavy-chain gene that compromise the protein’s contractile function even in heterozygous individuals. In such individuals, the heart tries to compensate by hypertrophy (enlargement), which often results in fatal heart arrhythmia (irregular beating). In addition to myosin heavy-chain defects, defects that result in cardiomyopathies have been traced to mutations in other components of the contractile machinery, including actin, myosin light chains, tropomyosin and troponin, and structural components such as titin (discussed below).