Cell Migration Coordinates Force Generation with Cell Adhesion and Membrane Recycling
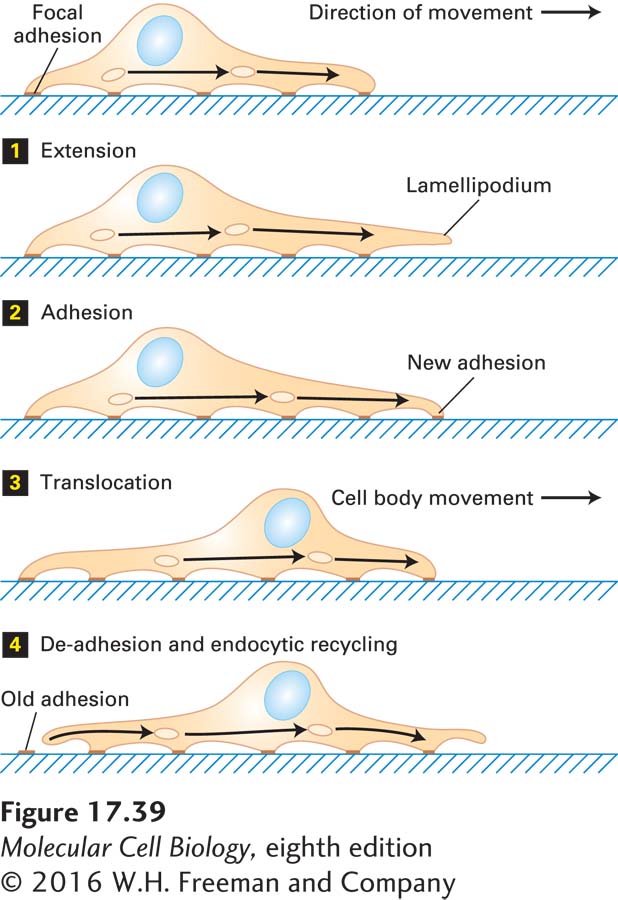
A moving fibroblast (connective tissue cell) displays a characteristic sequence of events: initial extension of a membrane protrusion, attachment to the substratum, forward flow of cytosol, and retraction of the rear of the cell (Figure 17-39). These events occur in an ordered pattern in a slowly moving cell such as a fibroblast, but in rapidly moving cells, such as macrophages, all of them are occurring simultaneously in a coordinated manner. We first consider the role of the actin cytoskeleton in cell movement, involving assembly at the leading edge as well as attachment to the substratum via stress fibers (Figure 17-40a, b), and then discuss the role of the endocytic cycle.
FIGURE 17-39Steps in cell locomotion. Movement begins with the extension of one or more lamellipodia from the leading edge of the cell 1; some lamellipodia adhere to the substratum by focal adhesions 2. Then the bulk of the cytoplasm in the cell body flows forward due to contraction at the rear of the cell 3. The trailing edge of the cell remains attached to the substratum until the tail eventually detaches and retracts into the cell body. During this cytoskeleton-based cycle, the endocytic cycle internalizes membrane and integrins at the rear of the cell and transports them to the front of the cell (arrows) for reuse in making new adhesions 4.
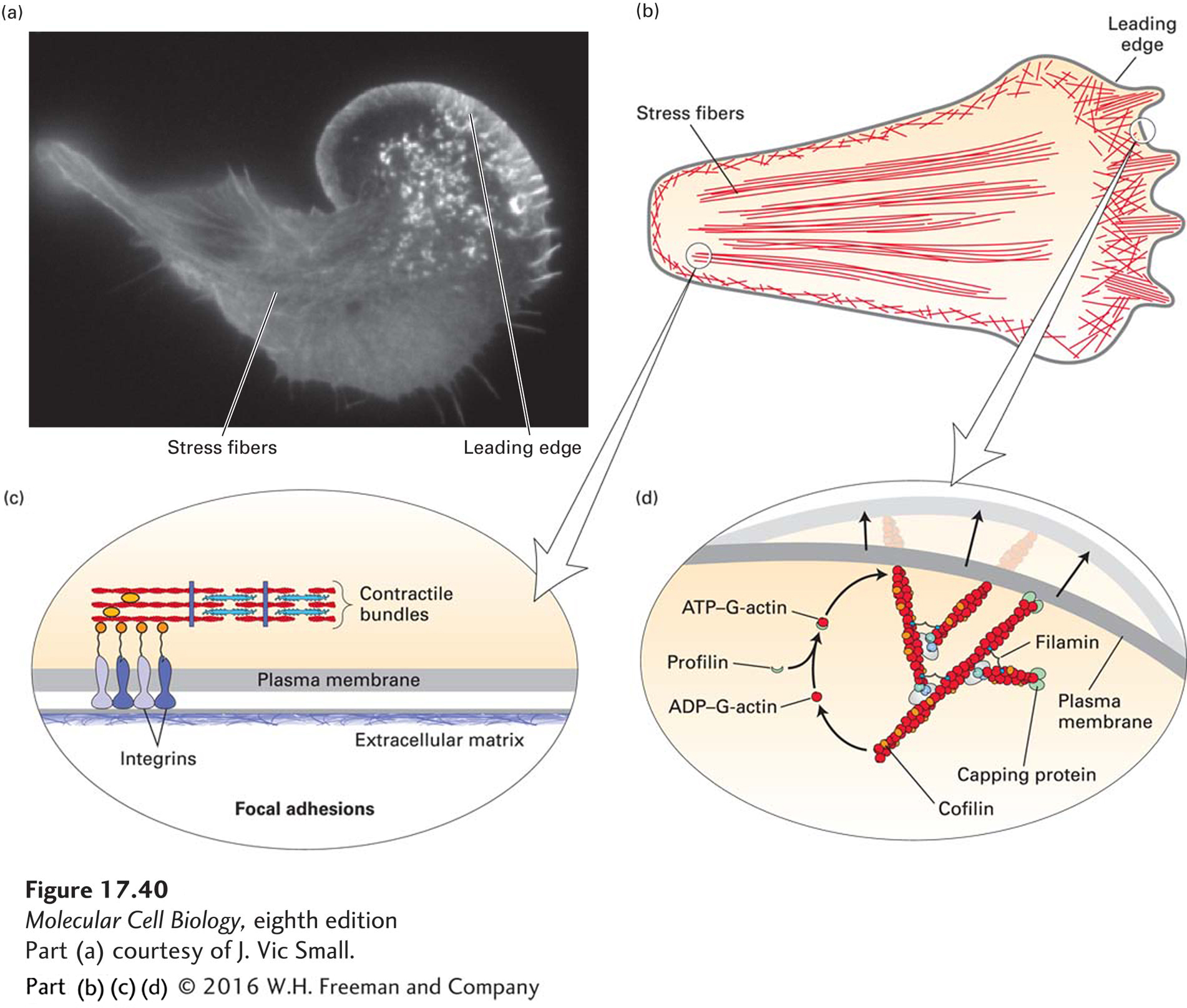
Membrane Extension The network of actin filaments at the leading edge is a type of cellular engine that pushes the membrane forward in a manner very similar to the propulsion of Listeria by actin polymerization (Figure 17-40d; for Listeria, see Figure 17-17c). At the membrane of the leading edge, actin is nucleated by the activated Arp2/3 complex, and filaments are elongated by assembly onto (+) ends adjacent to the plasma membrane. Because the actin network is fixed with respect to the substratum, the front membrane is pushed out as the filaments elongate. This process is very similar to the movement of Listeria, which “rides” on the polymerizing actin tail, which is also fixed within the cytoplasm. Actin turnover, and thus treadmilling, is mediated, as it is in the comet tails of Listeria, by the action of profilin and cofilin (see Figure 17-40d).
[Part (a) courtesy of J. Vic Small.]
FIGURE 17-40Actin-based structures involved in cell locomotion. (a) Localization of actin in a fibroblast expressing GFP-actin. (b) Diagram of the classes of microfilaments involved in cell migration. The network of actin filaments in the leading edge advances the cell forward. Contractile fibers in the cell cortex squeeze the cell body forward, and stress fibers terminating in focal adhesions also pull the bulk of the cell body up as the rear adhesions are released. (c) The structure of focal adhesions involves the attachment of the ends of stress fibers through integrins to the underlying extracellular matrix. Focal adhesions also contain many signaling molecules important for cell locomotion. (d) The dynamic actin network in the leading edge is nucleated by the Arp2/3 complex and employs the same set of factors that control assembly and disassembly of actin filaments in the Listeria tail (see Figure 17-17).
[Part (a) courtesy of J. Vic Small.]
Cell-Substratum Adhesions When the membrane has been extended and the actin network has been assembled, the plasma membrane becomes firmly attached to the substratum. Time-lapse microscopy shows that actin bundles in the leading edge become anchored to structures known as focal adhesions (Figure 17-40c). The attachment serves two purposes: it prevents the lamellipodium from retracting, and it attaches the cell to the substratum, allowing the cell to move forward. Given the importance of focal adhesions and their regulation during cell locomotion, it is not surprising that they have been found to be very rich in molecules involved in signal transduction pathways. Focal adhesions are discussed in more detail in Chapter 20, where we discuss cell-matrix interactions.
Page 812
The cell-adhesion molecules that mediate most cell-matrix interactions are membrane proteins called integrins. These proteins have an external domain that binds to specific components of the extracellular matrix, such as fibronectin and collagen, and a cytoplasmic domain that links them to the actin cytoskeleton (see Figure 17-40c and Chapter 20). The cell makes adhesions at the leading edge, and as the cell migrates forward, the adhesions eventually assume positions toward the rear.
Cell-Body Translocation After the forward adhesions have been made, the bulk contents of the cell body are translocated forward (see Figure 17-39, step 3). It is believed that the nucleus and the other organelles embedded in the cytoskeleton are moved forward by myosin II–dependent cortical contraction in the rear part of the cell, like toothpaste when the lower half of the tube is squeezed. Consistent with this model, myosin II is localized to the rear cell cortex.
Breaking Cell Attachments Finally, in the last step of movement (de-adhesion), the focal adhesions at the rear of the cell are broken, the integrins recycled, and the freed tail brought forward. In the light microscope, the tail is often seen to “snap” loose from its connections—perhaps by the contraction of stress fibers in the tail or by elastic tension—and it sometimes leaves a little bit of its membrane behind, still firmly attached to the substratum.
Cells cannot move if they are either too strongly attached or not attached to a surface. The ability of a cell to move corresponds to a balance between the mechanical forces generated by the cytoskeleton and the resisting forces generated by cell adhesions. This relationship can be demonstrated by measuring the rate of movement in cells that express varying levels of integrins. Such measurements show that the fastest migration occurs at an intermediate level of adhesion, with the rate of movement falling off at high and low levels of adhesion. Cell locomotion thus results from traction forces exerted by the cell on the underlying substratum.
Recycling of Membrane and Integrins by Endocytosis The dynamic changes in the actin cytoskeleton alone are not sufficient to drive cell migration; it is also dependent on endocytic recycling of membranes. The membrane needed during lamellipodium extension is provided by internal endosomes following their exocytosis. Adhesion molecules in focal adhesions at the rear of the cell are internalized as those adhesions are disassembled and transported by an endocytic cycle to the front to make new substratum attachments (Figure 17-39, step 4). This cycling of adhesion molecules in a migrating cell resembles the way a tank uses its treads to move forward. The movement of membrane internally through the cell also generates a rearward membrane flow across the surface of the cell. Indeed, this type of flow may contribute to the mechanics of cell locomotion, as it has recently been found that white blood cells can move in a liquid (“swim”) in the absence of attachment to a substratum, presumably as surface structures operating like paddles move backward across the cell surface.