Organelles in Axons Are Transported Along Microtubules in Both Directions
A neuron must constantly supply new materials—proteins and membranes—to its axon terminal to replenish those lost in the exocytosis of neurotransmitters at the junction (synapse) with another cell (see Chapter 22). Because proteins and membranes are primarily synthesized in the cell body, these materials must be transported down the axon, which can be as long as a meter in some neurons, to the synaptic region. This movement of materials is accomplished on microtubules, which are all oriented with their (+) ends toward the axon terminal (see Figure 18-5e).
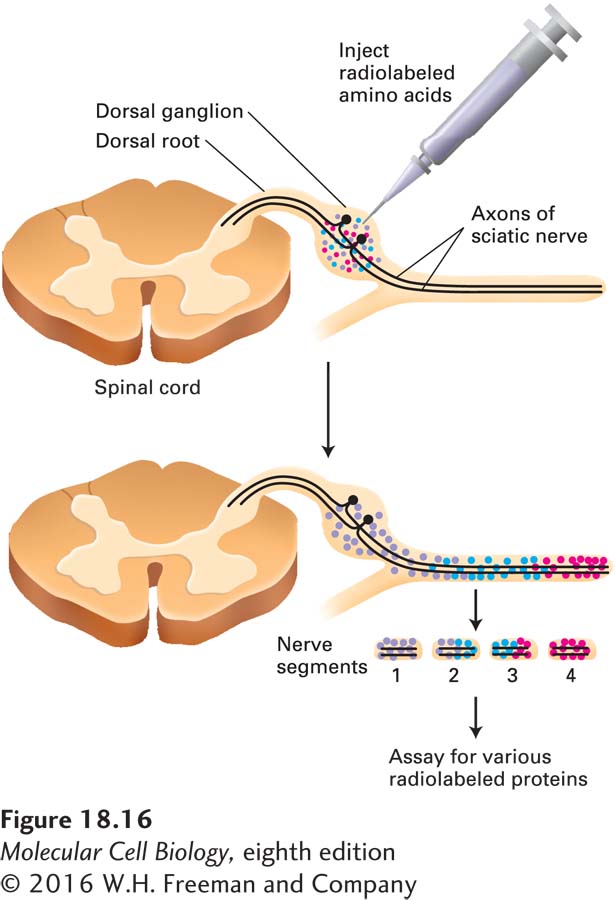
The results of classic pulse-chase experiments, in which radioactive amino acids were microinjected into the dorsal-root ganglia near the spinal cord to allow for their incorporation into proteins in spinal neurons, and the radioactivity was then tracked along the axons of those cells, showed that axonal transport occurs from the cell body down the axon. Other experiments showed that transport can also occur in the reverse direction, toward the cell body. Anterograde transport proceeds from the cell body to the axon terminal and is associated with axonal growth and the delivery of synaptic vesicles. In the opposite, retrograde, direction, “old” membranes from the axon terminal move along the axon rapidly toward the cell body, where they may be degraded in lysosomes. Findings from such experiments also revealed that different materials move at different speeds (Figure 18-16). The fastest-moving material, consisting of membrane-limited vesicles, has a velocity of about 3 mm/s, or 250 mm/day—requiring about 4 days to travel from a cell body in your back down an axon that terminates in your big toe. The slowest-moving material, comprising tubulin subunits and neurofilaments (the intermediate filaments found in neurons), moves only a fraction of a millimeter per day. Organelles such as mitochondria move down the axon at an intermediate rate.
EXPERIMENTAL FIGURE 18-16The rate of axonal transport in vivo can be determined by radiolabeling and gel electrophoresis. The cell bodies of neurons in the sciatic nerve are located in dorsal-root ganglia (near the spinal cord). Radioactive amino acids injected into these ganglia in experimental animals are incorporated into newly synthesized proteins, which are then transported down the axon to the synapse. Animals are sacrificed at various times after injection and the dissected sciatic nerve is cut into small segments to see how far radioactively labeled proteins have been transported; these proteins can be identified after gel electrophoresis and autoradiography. The red, blue, and purple dots represent groups of proteins that are transported down the axon at different rates, red most rapidly, purple least rapidly.
Page 834
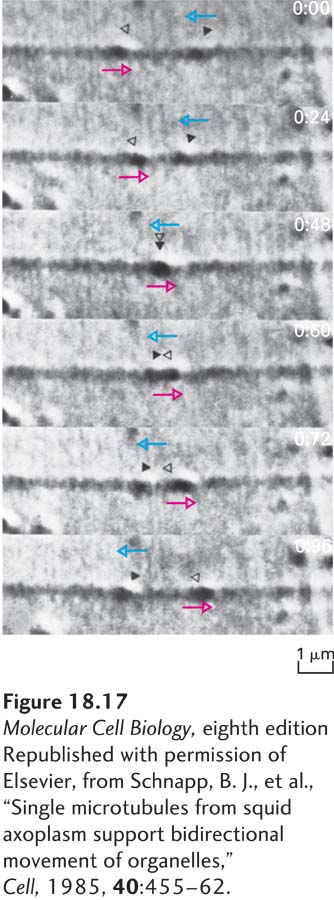
Neurobiologists have long made extensive use of the squid giant axon for studying organelle movement along microtubules. Involved in regulating the squid’s water propulsion system, the aptly named giant axon can be up to 1 mm in diameter, which is about 100 times wider than the average mammalian axon. Moreover, squeezing the axon like a tube of toothpaste results in the extrusion of the cytoplasm (also known as axoplasm), which can then be observed by video microscopy. The movement of vesicles along microtubules in this cell-free system requires ATP, its rate is similar to that of axonal transport in intact cells, and it can proceed in both the anterograde and the retrograde directions (Figure 18-17). Electron microscopy of the same region of squid giant axon cytoplasm revealed organelles attached to individual microtubules. These pioneering in vitro experiments established definitively that organelles move along individual microtubules and that their movement requires ATP.
[Republished with permission of Elsevier, from Schnapp, B. J., et al., “Single microtubules from squid axoplasm support bidirectional movement of organelles,” Cell, 1985, 40:455–62.]
EXPERIMENTAL FIGURE 18-17DIC microscopy demonstrates microtubule-based vesicle transport in vitro. Cytoplasm was squeezed from a squid giant axon with a roller onto a glass coverslip. After buffer containing ATP was added to the preparation, it was viewed by differential interference contrast (DIC) microscopy, and the images were recorded on videotape. In the sequential images shown, the two organelles indicated by open and solid triangles move in opposite directions (indicated by colored arrows) along the same filament, pass each other, and continue in their original directions. Elapsed time in seconds appears at the upper-right corner of each video frame.
[Republished with permission of Elsevier, from Schnapp, B. J., et al., “Single microtubules from squid axoplasm support bidirectional movement of organelles,” Cell, 1985, 40:455–62.]
Findings from experiments in which neurofilaments tagged with green fluorescent protein (GFP) were injected into cultured cells suggest that neurofilaments pause frequently as they move down an axon. Although the peak velocity of neurofilaments is similar to that of fast-moving vesicles, their numerous pauses lower the average rate of transport. These findings suggest that there is no fundamental difference between fast and slow axonal transport, although why neurofilament transport stops periodically is unknown.