Microtubule Walls Are Polarized Structures Built from αβ-Tubulin Dimers
Tubulin isolated in a pure and soluble form consists of two closely related subunits called α- and β-tubulin, each with a molecular weight of about 55,000 Da. Genomic analyses reveal that genes encoding both α- and β-tubulins are present in all eukaryotes, and that the number of genes has expanded considerably in multicellular organisms. For example, budding yeast has two genes specifying α-tubulin and one for β-tubulin, whereas the soil nematode Caenorhabditis elegans has nine genes encoding α-tubulin and six for β-tubulin. In addition to α- and β-tubulin, all eukaryotes also have genes specifying a third tubulin, γ-tubulin, which is involved in microtubule assembly, as we will see shortly. Additional isoforms of tubulin have also been discovered that are present only in organisms that possess cellular structures called centrioles and basal bodies, suggesting that these tubulin isoforms are important for those structures. As we’ll learn in this chapter, centrioles and basal bodies are specialized structures that some organisms use to nucleate and organize microtubule assembly.
Page 823
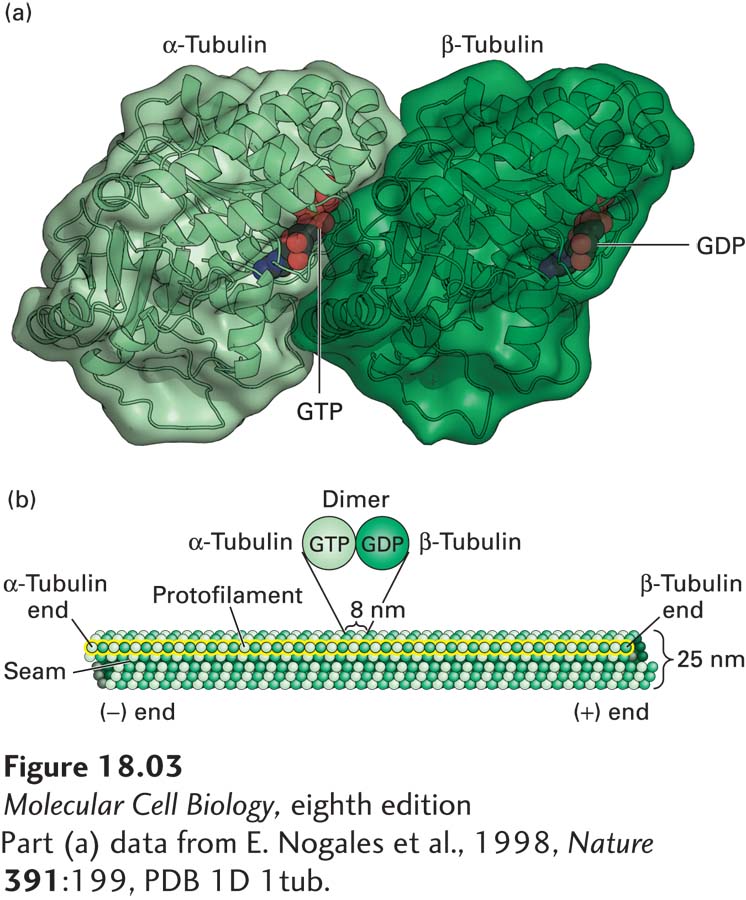
The α- and β-subunits of the tubulin dimer can each bind one molecule of GTP (Figure 18-3a). The GTP in the α-tubulin subunit is never hydrolyzed and is trapped by the interface between the α- and β-subunits. By contrast, the GTP-binding site on the β-subunit is at the surface of the dimer. GTP bound by the β-subunit can be hydrolyzed, and the resulting GDP can be exchanged for free GTP. Under appropriate conditions, soluble tubulin dimers can assemble into microtubules (Figure 18-3b). As we saw in Chapter 17 for the polymerization of actin, ATP–G-actin is preferentially added to one end of the filament, designated the (+) end because it is the end favored for assembly. Once incorporated into the filament, the bound ATP is hydrolyzed to ADP and Pi. In a similar manner, tubulin dimers in which the β-subunit has bound GTP are added preferentially to one end of the microtubule, also designated the (+) end. As we will see, the GTP is hydrolyzed once tubulin is incorporated into the microtubule, but in contrast to ATP hydrolysis in an actin filament, this GTP hydrolysis has dramatic effects on the behavior of the microtubule (+) end.
[Part (a) data from E. Nogales et al., 1998, Nature391:199, PDB 1D 1tub.]
FIGURE 18-3Structure of tubulin dimers and their organization into microtubules. (a) Ribbon diagram of the tubulin dimer. The GTP bound to the α-tubulin monomer is nonexchangeable, whereas the GDP bound to the β-tubulin monomer is exchangeable with free GTP. (b) The organization of tubulin subunits in a microtubule. The dimers are aligned end to end into protofilaments, which pack side by side to form the wall of the microtubule. The protofilaments are slightly staggered so that α-tubulin in one protofilament is in contact with α-tubulin in the neighboring protofilaments, except at the seam, where an α-subunit contacts a β-subunit. The microtubule displays a structural polarity in that subunits are added preferentially at the end where β-tubulin monomers are exposed. This end of the microtubule is known as the (+) end.
[Part (a) data from E. Nogales et al., 1998, Nature391:199, PDB 1D 1tub.]
A microtubule is composed of 13 laterally associated protofilaments, which form a tubule whose external diameter is about 25 nm (see Figure 18-3b). Each of the 13 protofilaments is a string of αβ-tubulin dimers, longitudinally arranged so that the subunits alternate down a protofilament, with each subunit type repeating every 8 nm. Because the αβ-tubulin dimers in a protofilament are all oriented in the same way, each protofilament has an α-subunit at one end and a β-subunit at the other—thus the protofilaments have an intrinsic polarity. In a microtubule, all the laterally associated protofilaments have the same polarity, so the microtubule also has an overall polarity. The end with exposed β-subunits is the (+) end, while the end with exposed α-subunits is the (−) end. In microtubules, the heterodimers in adjacent protofilaments are staggered slightly, forming tilted rows of α- and β-tubulin monomers in the microtubule wall. If you follow a row of β-subunits, for example, spiraling around a microtubule for one full turn, you will end up precisely three subunits up the protofilament, abutting an α-subunit. Thus all microtubules have a single longitudinal seam, where an α-subunit in one protofilament meets a β-subunit in the adjacent protofilament.
Page 824
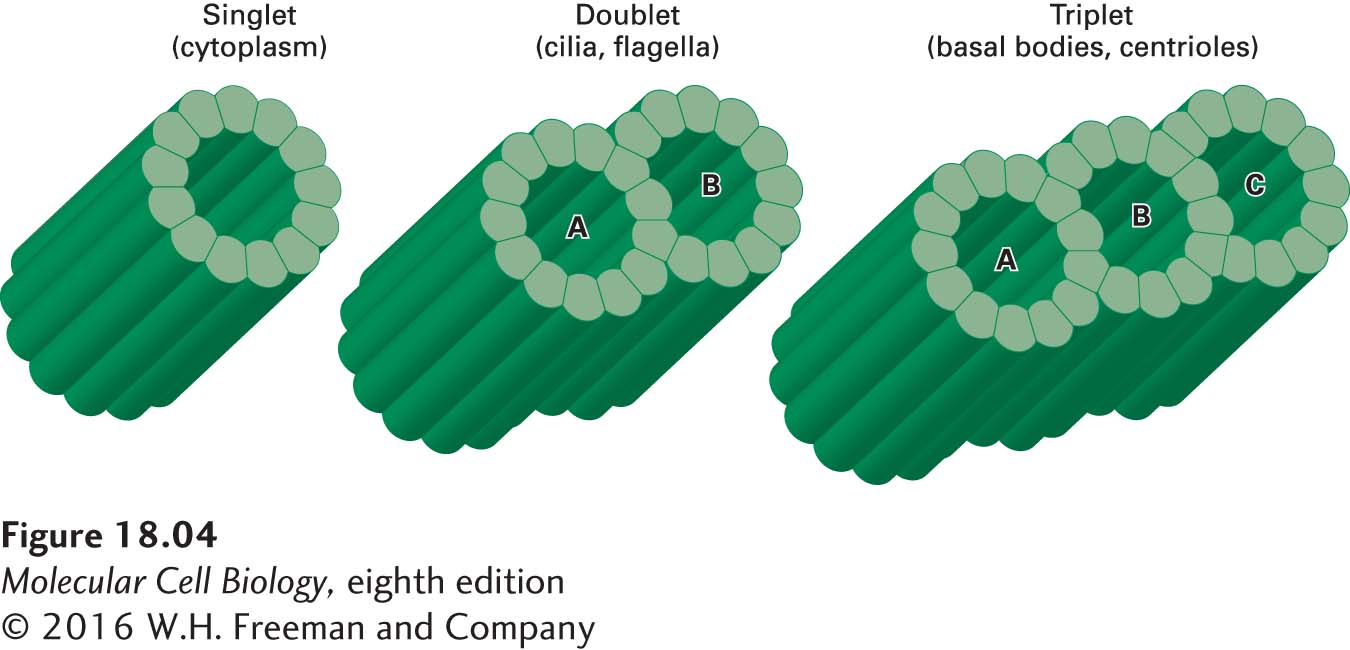
Most microtubules in a cell consist of a simple tube, a singlet microtubule, built from 13 protofilaments. In rare cases, singlet microtubules contain more or fewer protofilaments; for example, certain microtubules in the neurons of nematode worms contain 11 or 15 protofilaments. In addition to this simple singlet structure, doublet or triplet microtubules are found in specialized structures such as cilia and flagella (doublet microtubules) and centrioles and basal bodies (triplet microtubules), structures we will explore later in the chapter. Each doublet or triplet contains one complete 13-protofilament microtubule (called the A tubule) and one or two additional tubules (B and C) consisting of 10 protofilaments each (Figure 18-4).
FIGURE 18-4Singlet, doublet, and triplet microtubules. In cross section, a typical microtubule, a singlet, is a simple tube built from 13 protofilaments. In a doublet microtubule, an additional set of 10 protofilaments forms a second tubule (B) by fusing to the wall of a singlet (A) microtubule. Attachment of another 10 protofilaments to the (B) tubule of a doublet microtubule creates a (C) tubule and a triplet structure.