Replication at Each Origin Is Initiated Once and Only Once During the Cell Cycle
As discussed in Chapter 5, eukaryotic chromosomes are replicated from multiple replication origins. Initiation of replication from these origins occurs throughout S phase. However, no eukaryotic origin initiates replication more than once per S phase. Moreover, S phase continues until replication from multiple origins along the length of each chromosome has resulted in complete replication of the entire chromosome. These two factors ensure that the correct gene copy number is maintained as cells proliferate.
S phase CDKs play an essential role in the regulation of DNA replication. The kinases initiate DNA replication only at the G1–S phase transition and prevent re-initiation from origins that have already fired. We first discuss how initiation of DNA replication is controlled, and the role of S phase CDKs in the process, before turning to the mechanisms whereby these kinases prevent re-initiation.
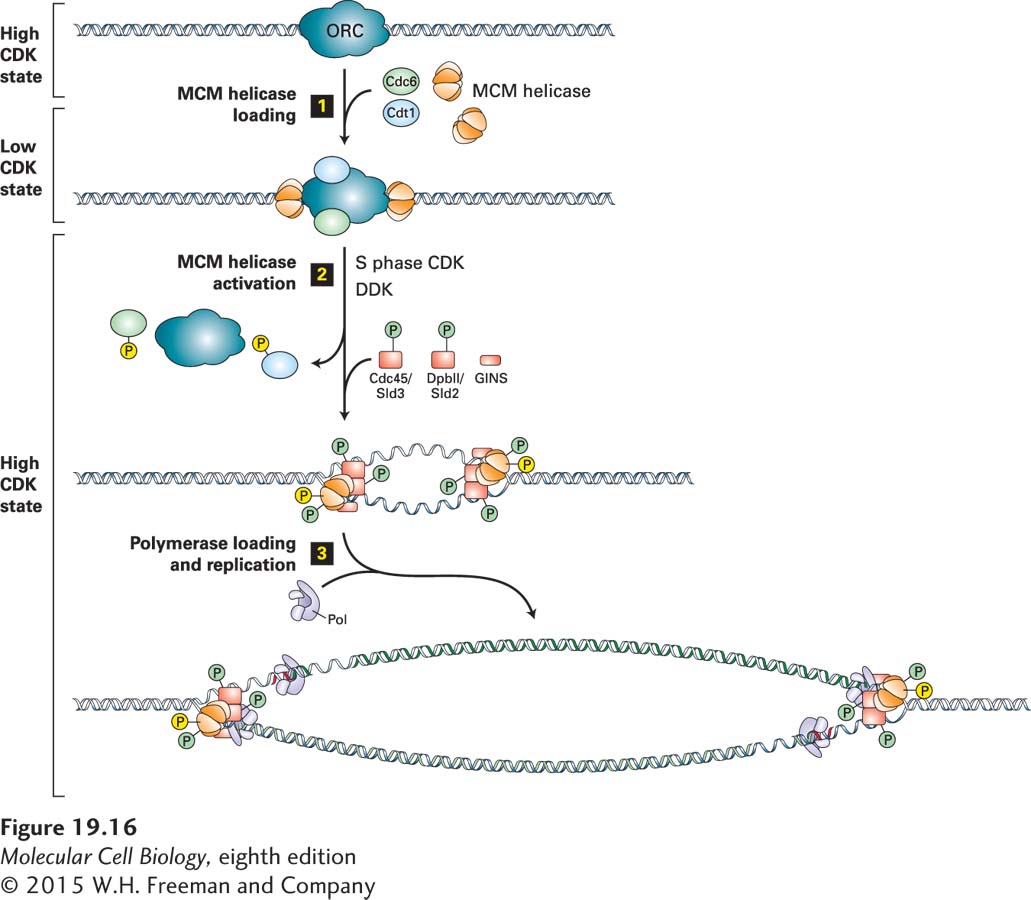
The mechanisms underlying the initiation of DNA replication are best understood in budding yeast, so we focus our discussion on this organism. However, it is important to note that the proteins and mechanisms controlling the initiation of DNA synthesis are essentially the same in all eukaryotic species. A protein complex known as the origin-recognition complex (ORC) is associated with all DNA replication origins. In budding yeast, replication origins contain an 11-bp conserved core sequence to which ORC binds. In multicellular organisms, DNA replication origins lack a recognizable consensus sequence. Instead, chromatin-associated factors target ORC to the DNA. ORC and two additional replication initiation factors, Cdc6 and Cdt1, associate with the ORC at origins during G1 to load the replicative helicases known as the MCM helicase complex onto DNA (Figure 19-16, step 1). The MCM helicases function to unwind the DNA during the initiation of DNA replication.
FIGURE 19-16The molecular mechanisms governing the initiation of DNA replication. Step 1: During exit from mitosis and early G1, when CDK activity is low, the MCM loading factors ORC, Cdc6, and Cdt1 load the replicative helicase, the MCM complex, onto DNA at replication origins. Step 2: Activation of S phase CDKs and DDK marks the onset of S phase. They phosphorylate the MCM helicase, Sld2, and Sld3 (depicted as green phosphorylation events), to facilitate the loading of MCM helicase activators—the Cdc45-Sld3 and GINS complexes—onto sites of replication initiation. Loading of these activators leads MCM helicases to unwind DNA. S phase CDKs also prevent reloading of MCM helicases by phosphorylating Cdc6 and Cdt1 (shown as yellow phosphorylation events), promoting their release from the replication origins and their degradation by SCF. S phase CDKs also phosphorylate MCM helicases, which leads to their export from the nucleus when the helicases disengage from the DNA when replication is complete. Step 3: DNA polymerases are recruited to origins, which leads to the initiation of DNA synthesis (see Figure 5-30).
To ensure that origins fire only once at the beginning of S phase, the loading of MCM helicase complex and its activation occur at two opposing phosphorylation states. The MCM helicases can be loaded onto the DNA only in a state of low CDK activity that occurs when CDKs are inactivated during exit from mitosis and during early G1. In other words, MCM helicases are loaded onto DNA when they are nonphosphorylated. In contrast, activation of the MCM helicases and the recruitment of DNA polymerases to the unwound origin DNA are triggered by S phase CDKs. Recall that S phase CDKs become active only when G1/S phase CDK levels reach their peak and the CKIs of S phase CDKs are destroyed. It is then that phosphorylation by S phase CDKs and a second heterodimeric protein kinase, DDK, activates the MCM helicases and recruits DNA polymerases to the sites of replication initiation (Figure 19-16, steps 2 and 3).
So how do S phase CDKs and DDK collaborate to initiate DNA replication? ORC and the two other initiation factors, Cdc6 and Cdt1, recruit the MCM helicases to sites of replication initiation during G1, when CDK activity is low (see Figure 19-16, step 1). When DDK and S phase CDKs are activated in late G1, DDK phosphorylates two subunits of the MCM helicase. The S phase CDKs phosphorylate two proteins called Sld2 and Sld3. These phosphorylation events (shown in green in Figure 19-16, steps 2 and 3) have an activating effect, promoting the recruitment of MCM helicase activators to sites of replication initiation. The helicase activators are called the Cdc45-Sld3 complex and the GINS complex. Exactly how they promote activation of the MCM helicases is not yet clear. In addition to activating the MCM helicase to unwind DNA, they recruit polymerases to the DNA: polymerase ε to synthesize the leading strand and polymerase δ to synthesize the lagging strand (see Figure 19-16, step 3). The replication machinery then initiates DNA synthesis.
S phase CDKs are not only essential for initiating DNA replication, but are also responsible for ensuring that each origin fires only once during S phase. Re-firing of origins during S phase is prevented by phosphorylation of several components of the MCM helicase loading machinery and the MCM helicase complex itself. To distinguish these phosphorylation events from the ones required for the initiation of DNA replication, they are depicted in yellow in Figure 19-16. Concomitant with activation of the MCM helicases, Cdc6 and Cdt1 dissociate from the sites of DNA replication initiation. Once they have done so, their phosphorylation leads to their degradation by the SCF ubiquitin-protein ligase. Phosphorylation of the MCM helicases leads to the export of these proteins from the nucleus after they dissociate from the DNA on completion of DNA replication. Thus only after CDK activity is lowered by APC/CCdh1 during exit from mitosis can the MCM helicases be reloaded onto DNA. As a result, helicase loading is restricted to late stages of mitosis and early G1 (see Figure 19-16, step 1).
The general mechanisms governing the initiation of DNA replication in metazoan cells parallel those in S. cerevisiae, although small differences are found in vertebrates. The helicases are loaded in G1, when CDK activity is low. Phosphorylation of MCM helicase activators by G1/S phase CDKs and S phase CDKs activates the helicases and promotes polymerase loading. As in yeast, phosphorylation of MCM helicase loading factors prevents reloading of MCM helicases until the cell passes through mitosis, thereby ensuring that replication from each origin occurs only once during each cell cycle. Here Cdt1 phosphorylation by multiple protein kinases, including CDKs, is especially critical for preventing reloading of the MCM helicases. In addition, a small protein, geminin, contributes to the inhibition of re-initiation at origins until cells complete a full cell cycle. Geminin is expressed in late G1; it binds to and inhibits MCM helicase loading factors as they are released from origins once DNA replication is initiated during S phase (see Figure 19-16, step 2). Geminin contains a destruction box at its N-terminus that is recognized by APC/CCdh1, causing it to be ubiquitinylated in late anaphase and degraded by proteasomes. Its degradation frees the MCM helicase loading factors, which are also dephosphorylated as CDK activity declines, to bind to ORC on replication origins and load MCM helicases during the following G1 phase.