Mitotic CDK Inactivation Triggers Exit from Mitosis
Anaphase spindle elongation and the events associated with exit from mitosis—mitotic spindle disassembly, chromosome decondensation, and nuclear envelope re-formation—are brought about by the dephosphorylation of CDK substrates. In other words, exit from mitosis can be viewed as a reversal of entry into mitosis. The phosphorylation events that triggered the different mitotic events need to be undone for the cell to revert to the G1 state.
Dephosphorylation of mitotic CDK substrates is caused by the inactivation of mitotic CDKs. In most organisms, mitotic CDK inactivation is triggered by APC/CCdc20-mediated degradation of mitotic cyclins. As mitotic CDKs activate APC/CCdc20, they initiate their own demise. In budding yeast, only about 50 percent of mitotic cyclins are degraded by APC/CCdc20. As we will see in Section 19.7, a pool of mitotic cyclins is protected from APC/CCdc20 to allow for enough time to position the mitotic spindle accurately within the cell. How a fraction of mitotic cyclins is protected from APC/CCdc20 is not known, but it is clear that a second mitotic CDK-inactivating step is needed for exit from mitosis to occur. The conserved protein phosphatase Cdc14 brings about this second step in mitotic CDK inhibition.
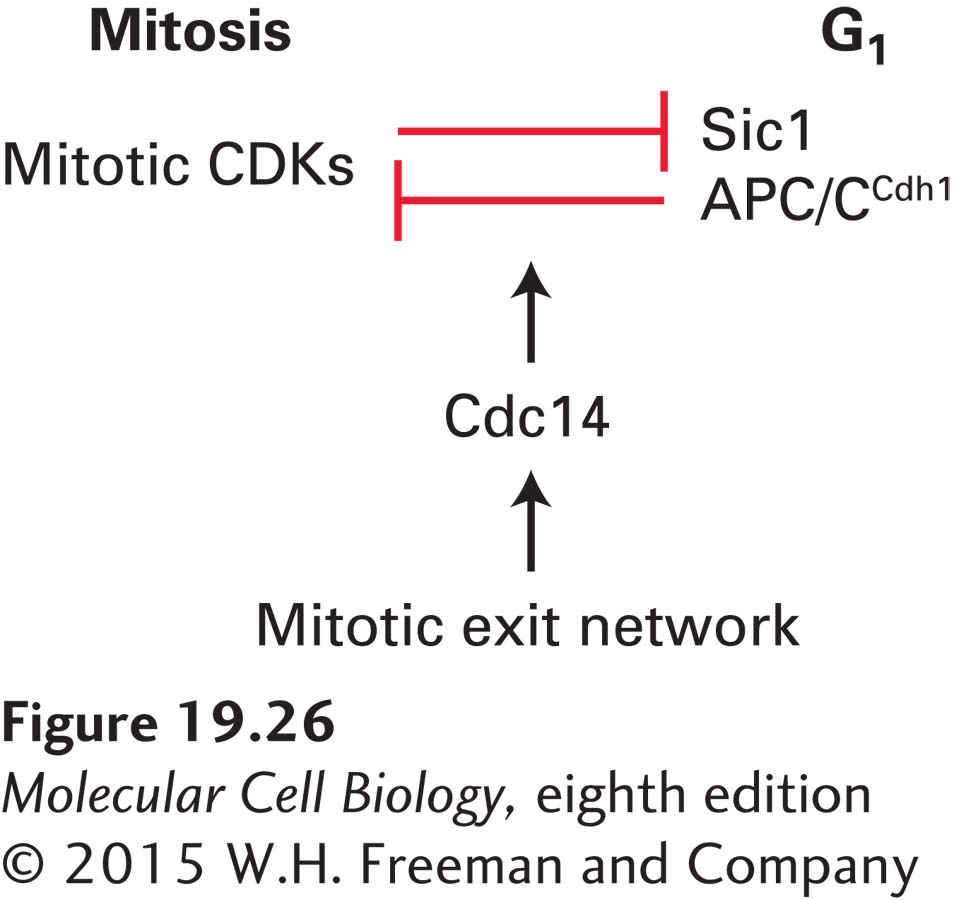
FIGURE 19-26The protein phosphatase Cdc14 triggers exit from mitosis in budding yeast. During mitosis, mitotic CDK activity inhibits its inhibitors, APC/CCdh1 and Sic1. During G1, APC/CCdh1 and Sic1 inhibit mitotic CDKs. During exit from mitosis, the protein phosphatase Cdc14 throws the switch between these two antagonistic states. The mitotic exit network activates the phosphatase during anaphase, allowing it to dephosphorylate APC/CCdh1, thereby activating it. The phosphatase also promotes the accumulation of Sic1. In addition, Cdc14 dephosphorylates the many mitotic CDK substrates, which leads to rapid exit from mitosis.
In budding yeast, complete inactivation of mitotic CDKs requires the destruction of mitotic cyclins by APC/CCdh1 and the accumulation of the CDK inhibitor Sic1, which—recall—holds S phase CDKs in check until cells enter the cell cycle. Both APC/CCdh1 and Sic1 are inhibited by mitotic CDKs. Conversely, APC/CCdh1 and Sic1 inhibit mitotic CDKs (Figure 19-26). The protein phosphatase Cdc14 throws the switch between these two mutually antagonistic states during anaphase. Cdc14 is kept inactive during most of the cell cycle, but is activated during anaphase by a GTPase signaling pathway known as the mitotic exit network. This signaling cascade, as we will see in Section 19.7, is responsive to spindle position and becomes active only in anaphase when the anaphase spindle is properly positioned within the cell. Once activated during anaphase, Cdc14 dephosphorylates APC/CCdh1 and Sic1 to promote mitotic cyclin degradation and mitotic CDK inactivation, respectively. This process leads to exit from mitosis.
Phosphatase activity is also essential for exit from mitosis in vertebrates. Simple inactivation of mitotic CDKs is not sufficient to trigger a timely exit from mitosis. It is not yet clear which phosphatase dephosphorylates CDK substrates to reset the cell to the G1 stage. Both protein phosphatase 1 and protein phosphatase 2A have been implicated in the process.
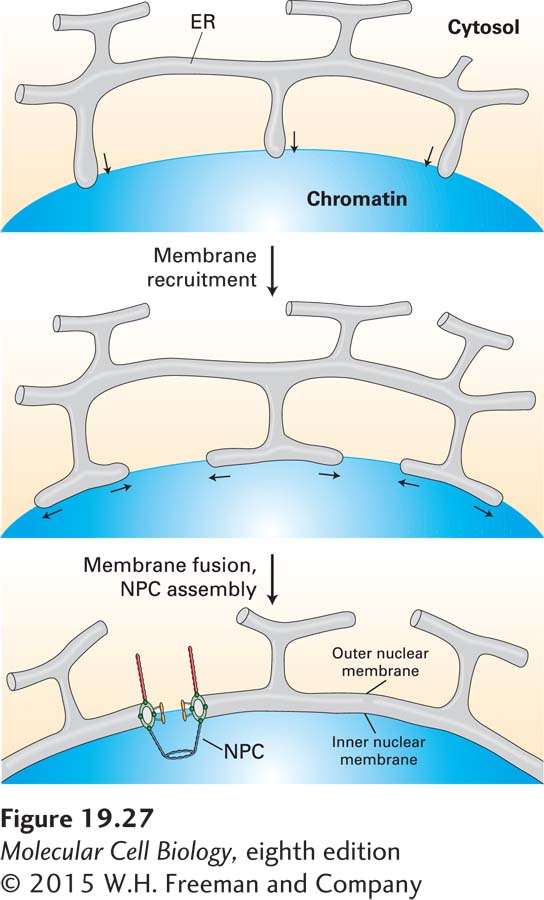
Ultimately, reversal of mitotic CDK phosphorylation changes the activities of many proteins back to their interphase states. Dephosphorylation of condensins, histone H1, and other chromatin-associated proteins leads to the decondensation of mitotic chromosomes in telophase. The targets of CDKs whose dephosphorylation is important for mitotic spindle disassembly are not known, but it is likely that multiple proteins are targets. More is known about how the nuclear envelope re-forms. Dephosphorylated inner nuclear membrane proteins are thought to bind to chromatin once again. As a result, multiple projections of regions of the ER membrane containing these proteins are thought to associate with the surfaces of the decondensing chromosomes and then fuse with one another, directed by an unknown mechanism to form a continuous double membrane around each chromosome (Figure 19-27). Dephosphorylation of nuclear pore subcomplexes allows them to reassemble into complete NPCs traversing the inner and outer membranes soon after fusion of the ER projections. Ran-GTP, required for driving most nuclear import and export (see Chapter 13), stimulates both fusion of the ER projections to form daughter nuclear envelopes and assembly of NPCs (see Figure 19-27). The Ran-GTP concentration is highest in the microvicinity of the decondensing chromosomes because the Ran-guanine nucleotide-exchange factor (Ran-GEF) is bound to chromatin. Consequently, membrane fusion is stimulated at the surfaces of decondensing chromosomes. Sheets of nuclear membrane with inserted NPCs then fuse with one another to form one nuclear membrane around all chromosomes.
Page 903
FIGURE 19-27Model for reassembly of the nuclear envelope during telophase. Extensions of the endoplasmic reticulum (ER) associate with each decondensing chromosome and then fuse with one another, forming a double membrane around the chromosome. Dephosphorylated nuclear pore subcomplexes reassemble into nuclear pores, forming individual mini-nuclei called karyomeres. The enclosed chromosome further decondenses, and subsequent fusion of the nuclear envelopes of all the karyomeres at each spindle pole forms a single nucleus containing a full set of chromosomes. NPC, nuclear pore complex. See B. Burke and J. Ellenberg, 2002, Nature Rev. Mol. Cell Biol.3:487.