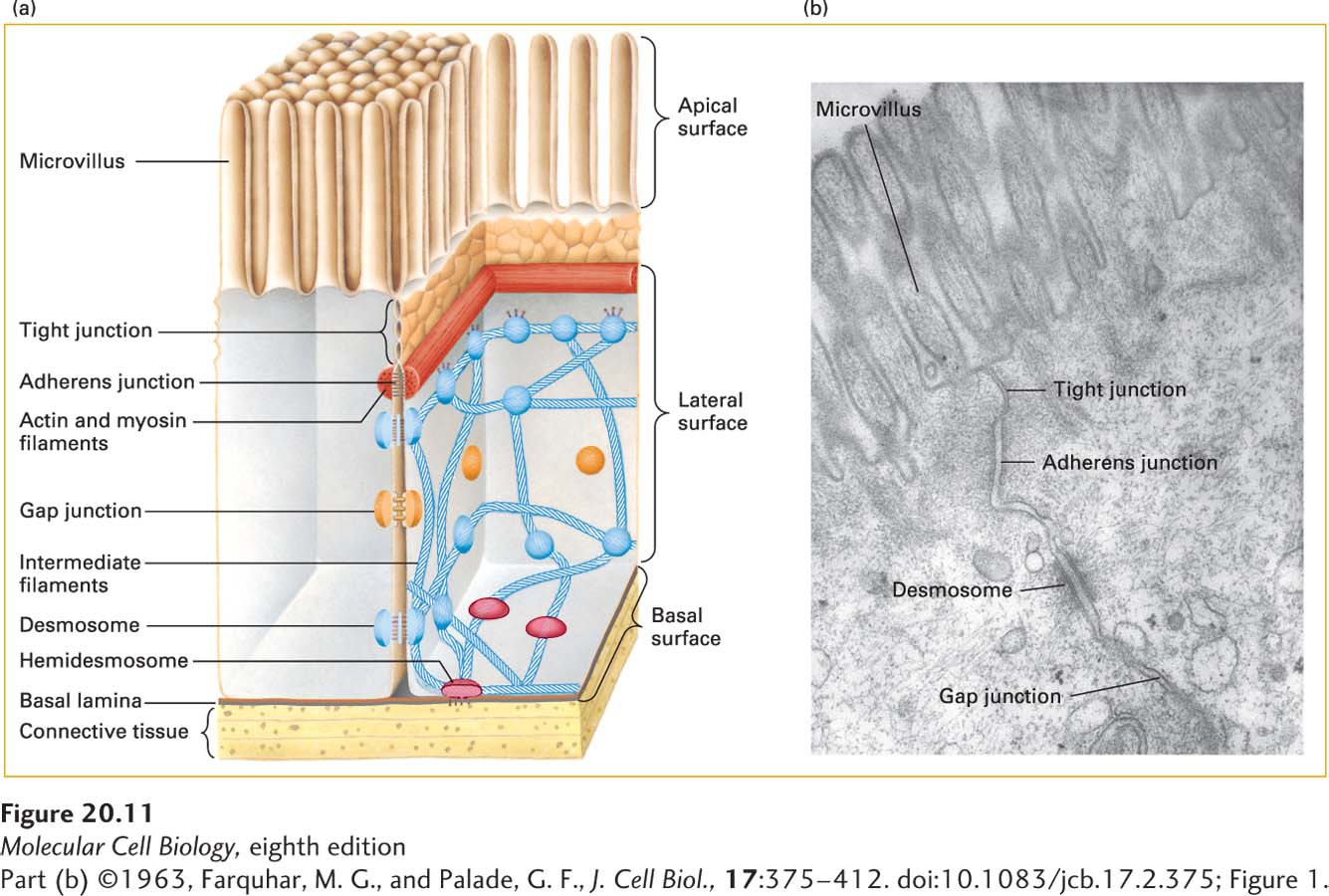
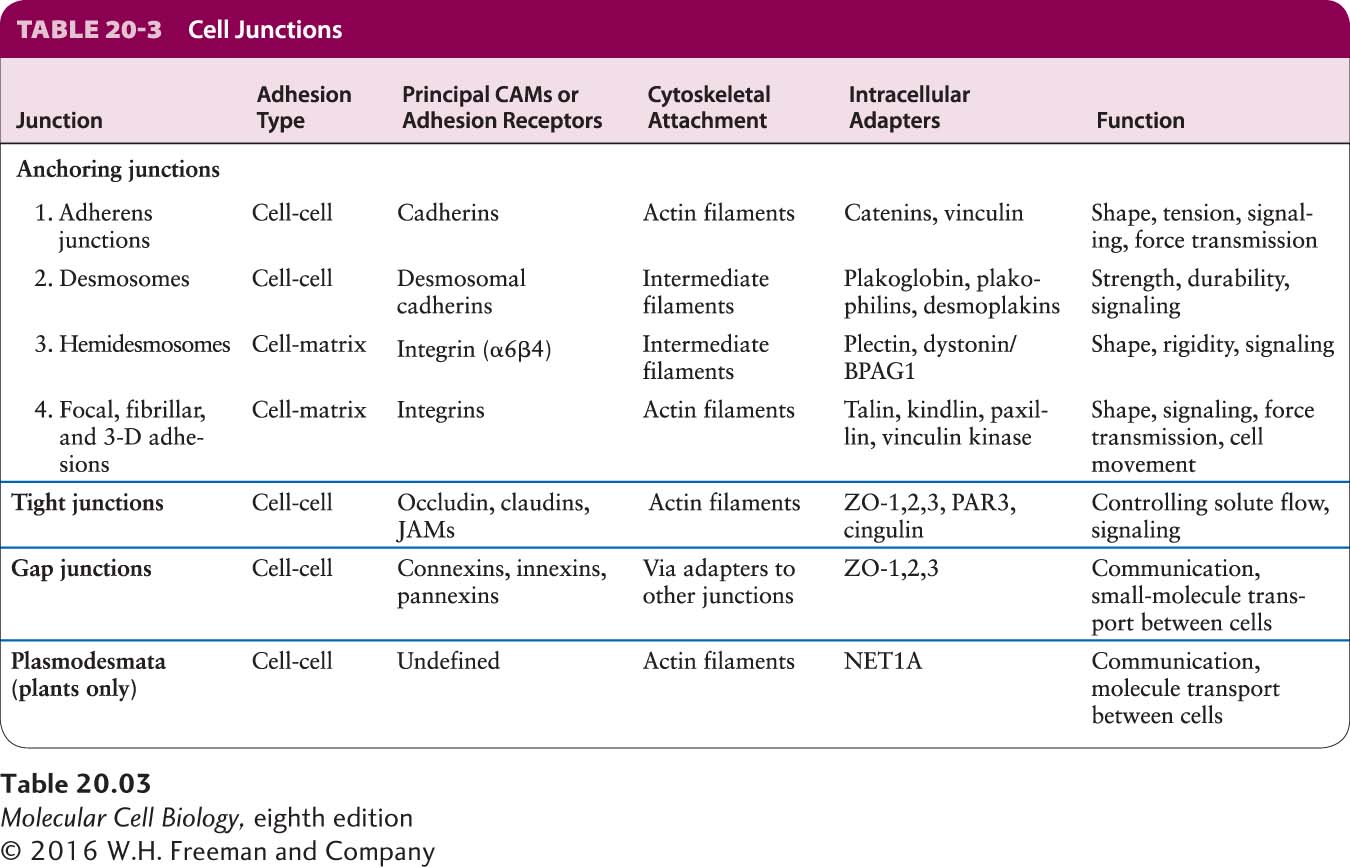
Three Types of Junctions Mediate Many Cell-Cell and Cell-ECM Interactions
All epithelial cells in a sheet are connected to one another and to the ECM by specialized junctions. Although hundreds of individual dispersed adhesion molecule–mediated interactions are sufficient to cause cells to adhere, the clustered groups of adhesion molecules at cell junctions play special roles in imparting strength and rigidity to a tissue, transmitting information between the extracellular and the intracellular space, controlling the passage of ions and molecules across cell layers, and serving as conduits for the movement of ions and molecules from the cytoplasm of one cell to that of its immediate neighbor. Particularly important to epithelia is the formation of junctions that help form tight seals between the cells and thus allow the epithelial sheet to serve as a barrier to the flow of molecules from one side of the sheet to the other.
Three major classes of animal-cell junctions are prominent features of simple columnar epithelia (Figure 20-11 and Table 20-3): anchoring junctions, tight junctions, and gap junctions. Anchoring junctions and tight junctions perform the key task of holding the tissue together. As we shall see, tight junctions also control the flow of solutes through the extracellular spaces between the cells forming an epithelial sheet. Tight junctions are found primarily in epithelial cells, whereas anchoring junctions can be seen in both epithelial and nonepithelial cells. Anchoring junctions and tight junctions in epithelia are organized into three parts: (1) adhesive proteins in the plasma membrane that connect one cell to another cell on the lateral surfaces (CAMs) or to the ECM on the basal surfaces (adhesion receptors); (2) adapter proteins, which connect the CAMs or adhesion receptors to cytoskeletal filaments and signaling molecules; and (3) the cytoskeletal filaments themselves.
FIGURE 20-11Principal types of cell junctions connecting the columnar epithelial cells lining the small intestine. (a) Schematic cutaway drawing of intestinal epithelial cells. The basal surface of the cells rests on a basal lamina, and the apical surface is packed with fingerlike microvilli that project into the intestinal lumen. Tight junctions, lying just under the microvilli, prevent the diffusion of many substances between the intestinal lumen and internal body fluids (such as the blood) via the extracellular space between cells. Gap junctions allow the movement of small molecules and ions between the cytosols of adjacent cells. The remaining three types of junctions—adherens junctions, desmosomes, and hemidesmosomes—are critical to cell-cell and cell-matrix adhesion and signaling. (b) Electron micrograph of a thin section of epithelial cells in the rat intestine, showing the relative locations of the different junctions.
The third class of junctions, gap junctions, permits the rapid diffusion of small, water-soluble molecules between the cytoplasms of adjacent cells. Along with anchoring and tight junctions, gap junctions help a cell communicate with its environment. However, they are structurally very different from anchoring junctions and tight junctions and do not play a key role in strengthening cell-cell and cell-ECM adhesions. Found in both epithelial and nonepithelial cells, gap junctions resemble the distinct cell junctions in plants called plasmodesmata, which we discuss in Section 20.6.
Page 933
Four types of anchoring junctions are present in cells. Two participate in cell-cell adhesion and two participate in cell-matrix adhesion. Adherens junctions connect the lateral membranes of adjacent epithelial cells and are usually located near the apical surface, just below the tight junctions (see Figure 20-11). A circumferential belt of actin and myosin filaments in a complex with the adherens junctions functions as a tension cable that can internally brace the cell and thereby control its shape. Epithelial and some other types of cells, such as smooth muscle and heart cells, are also bound tightly together by desmosomes, snap-like points of contact sometimes called spot desmosomes. Hemidesmosomes, found mainly on the basal surface of epithelial cells, and focal contacts (also called focal adhesions) anchor an epithelium to components of the underlying ECM, much like nails holding down a carpet. Adherens junctions, desmosomes, and focal adhesions are found in many different types of cells; hemidesmosomes appear to be restricted to epithelial cells.
Bundles of intermediate filaments running parallel to the cell surface or through the cell connect desmosomes and hemidesmosomes, imparting shape and rigidity to the cell, as do actin filaments that connect the cytoskeleton with focal contacts and adherens junctions. The close interaction between these junctions and the cytoskeleton helps transmit shear forces from one region of a cell layer to the epithelium as a whole, providing strength and rigidity to the entire epithelial cell layer. Desmosomes and hemidesmosomes are especially important in maintaining the integrity of skin epithelia. As a consequence, mutations that interfere with hemidesmosomal anchoring in the skin can lead to a condition in which the epithelium becomes detached from its underlying matrix and extracellular fluid accumulates at the basolateral surface, forcing the skin to balloon outward, forming a blister.