Proteoglycans and Their Constituent GAGs Play Diverse Roles in the ECM
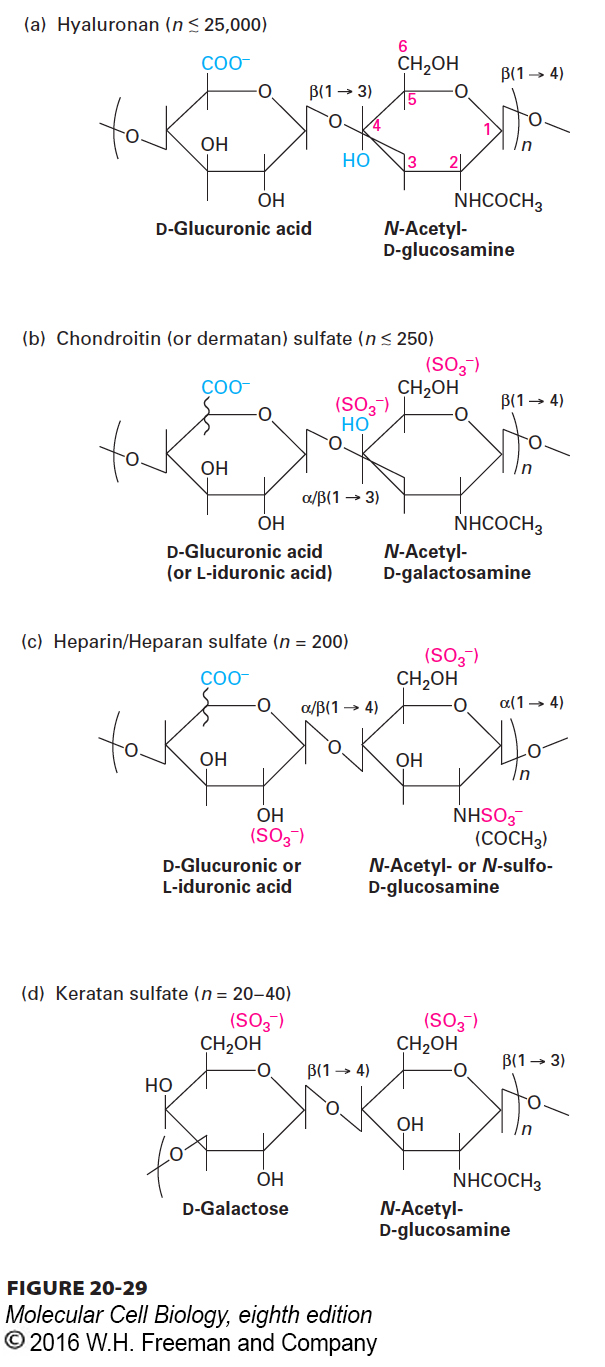
As we saw with perlecan in the basal lamina, proteoglycans play an important role in cell-ECM adhesion. Proteoglycans are a subset of secreted or cell-surface glycoproteins containing covalently linked, specialized polysaccharide chains called glycosaminoglycans (GAGs). GAGs are long linear polymers of specific repeating disaccharides. Usually one sugar is either a uronic acid (D-glucuronic acid or L-iduronic acid) or D-galactose; the other sugar is N-acetylglucosamine or N-acetylgalactosamine (Figure 20-29). One or both of the sugars contain at least one anionic group (carboxylate or sulfate). Thus each GAG chain bears many negative charges. GAGs are classified into several major types based on the nature of the repeating disaccharide unit: heparan sulfate, chondroitin sulfate, dermatan sulfate, keratan sulfate, and hyaluronan (Figure 20-29). A hypersulfated form of heparan sulfate called heparin, produced mostly by mast cells, plays a key role in allergic reactions. It is also used medically as an anticlotting drug because of its ability to activate a natural clotting inhibitor called antithrombin III.
FIGURE 20-29The repeating disaccharides of glycosaminoglycans (GAGs). Each of the four classes of GAGs is formed by polymerization of monomeric units into repeats of a particular disaccharide and subsequent modifications, including addition of sulfate groups and inversion (epimerization) of the carboxyl group on carbon 5 of D-glucuronic acid to yield L-iduronic acid. The squiggly lines represent covalent bonds that are oriented either above (D-glucuronic acid) or below (L-iduronic acid) the ring. Heparin is generated by hypersulfation of heparan sulfate, whereas hyaluronan is nonsulfated.
Page 954
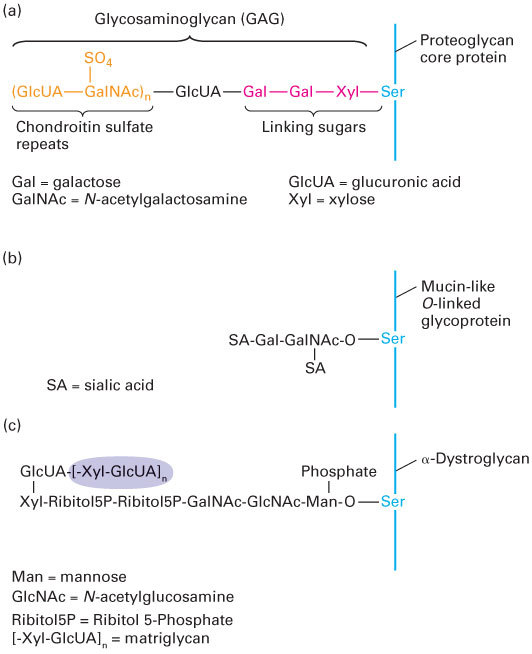
With the exception of hyaluronan, which is discussed below, all the major GAGs occur naturally as components of proteoglycans. Like other secreted and transmembrane glycoproteins, proteoglycan core proteins are synthesized in the endoplasmic reticulum, and the GAG chains are assembled on and covalently attached to these cores in the Golgi complex. To generate heparan or chondroitin sulfate chains, a three-sugar “linker” is first attached to the hydroxyl side chains of certain serine residues in a core protein; thus these GAGs are O-linked oligosaccharides (Figure 20-30). In contrast, the linkers for the addition of keratan sulfate chains are oligosaccharide chains attached to asparagine residues; such N-linked oligosaccharides are present in many glycoproteins (see Chapter 14), although only a subset carry GAG chains. All GAG chains are elongated by the alternating addition of sugar monomers to form the disaccharide repeats characteristic of a particular GAG; the chains are often modified subsequently by the covalent linkage of small molecules such as sulfate. The mechanisms responsible for determining which proteins are modified with GAGs, the sequence of disaccharides to be added, the sites to be sulfated, and the lengths of the GAG chains are unknown. The ratio of polysaccharide to protein in all proteoglycans is much higher than that in most other glycoproteins.
FIGURE 20-30Hydroxyl (O-) linked oligosaccharides. (a) Synthesis of a glycosaminoglycan (GAG), in this case chondroitin sulfate, is initiated by transfer of a xylose (Xyl) residue to a serine residue in the core protein, most likely in the Golgi complex, followed by sequential addition of two galactose (Gal) residues. Glucuronic acid (GlcUA) and N-acetylgalactosamine (GalNAc) residues are then added sequentially to these linking sugars and some of the GalNAc monomers are sulfated, forming the chondroitin sulfate chain. Heparan sulfate chains are connected to core proteins by the same three-sugar linker. Keratan sulfate GAGs are covalently attached to proteins via N-linked rather than O-linked connections. (b) Mucin-like O-linked chains are covalently bound to glycoproteins via an N-acetylgalactosamine (GalNAc) monosaccharide to which are covalently attached a variety of other sugars, often including sialic acid (SA). (c) Certain specialized O-linked oligosaccharides, such as those found in the adhesion receptor dystroglycan, are bound to proteins via mannose (Man) monosaccharides. The attachment of matriglycan, a polymer of the GlcUA-Xyl disaccharide (shaded), to the mannose via a phosphate and an additional unknown linkage (?) provides a binding site on dystroglycan for ECM molecules, such as laminin and perlecan.
Function of GAG Chain Modifications Similar to the sequence of amino acids in proteins, the arrangement of the sugar residues in GAG chains and the modification of specific sugars in those chains can determine their function and that of the proteoglycans that contain them. For example, groupings of certain modified sugars in the GAG chains of heparin sulfate proteoglycans can control the binding of growth factors to certain receptors or the activities of proteins in the blood-clotting cascade.
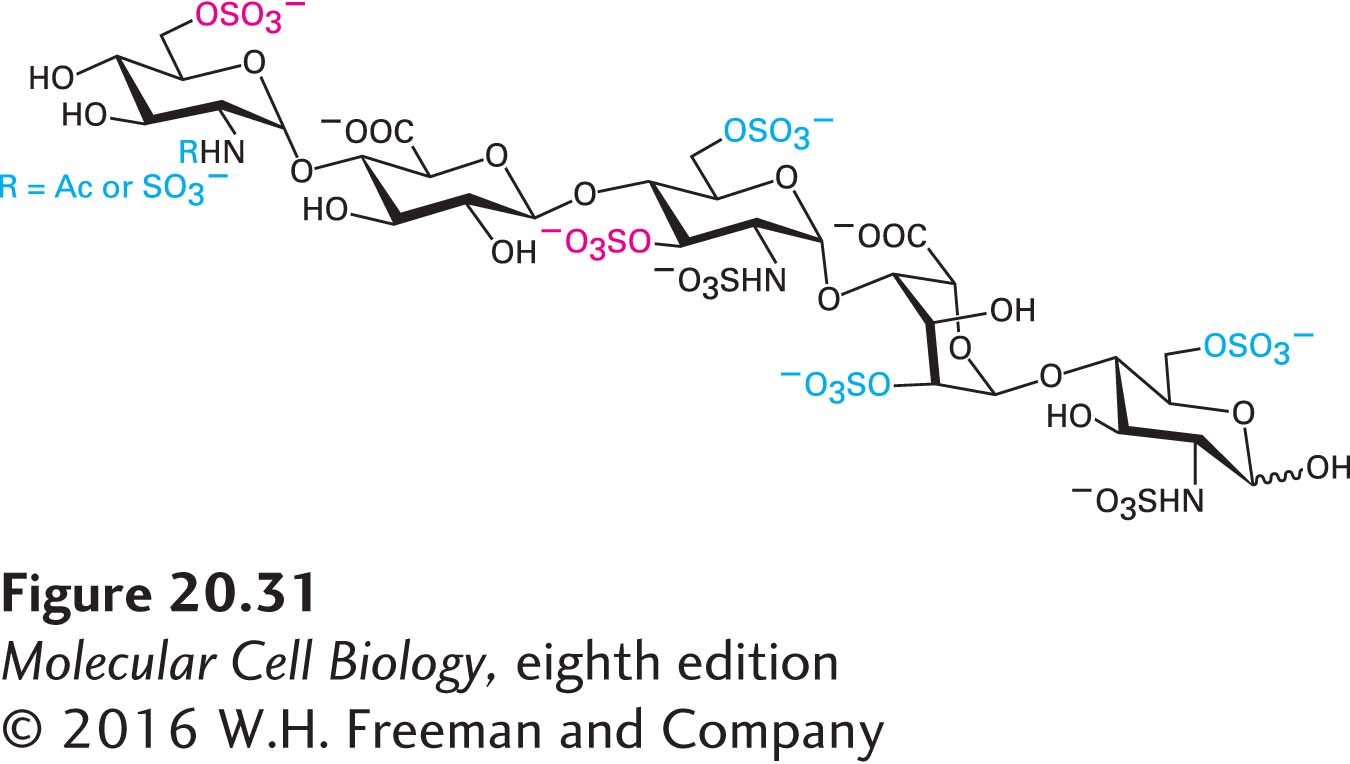
In the past, the chemical and structural complexity of proteoglycans posed a daunting barrier to analyzing and understanding their structures and their many diverse functions. In recent years, investigators employing classic and state-of-the-art biochemical techniques, mass spectrometry, and genetics have begun to elucidate the detailed structures and functions of these ubiquitous ECM molecules. The results of ongoing studies suggest that sets of sugar-residue sequences containing common modifications, rather than single unique sequences, are responsible for specifying distinct GAG functions. A case in point is a set of five-residue (pentasaccharide) sequences found in a subset of heparin GAGs that controls the activity of antithrombin III (ATIII), an inhibitor of the key blood-clotting protease thrombin. When these pentasaccharide sequences in heparin are sulfated at two specific positions (Figure 20-31), heparin can activate ATIII, thereby inhibiting clot formation. Several other sulfates can be present in the active pentasaccharide in various combinations, but they are not essential for the anticlotting activity of heparin. The rationale for generating sets of similar active sequences rather than a single unique sequence is not well understood.
FIGURE 20-31The pentasaccharide GAG sequence that regulates the activity of antithrombin III (ATIII). Sets of modified five-residue sequences in the much longer GAG called heparin with the composition shown here bind to ATIII and activate it, thereby inhibiting blood clotting. The sulfate groups in red type are essential for this heparin function; the modifications in blue type may be present but are not essential. Other sets of modified GAG sequences are thought to regulate the activity of other target proteins.
Page 955
Diversity of Proteoglycans The proteoglycans constitute a remarkably diverse group of molecules that are abundant in the ECM of all animal tissues and are also expressed on the cell surface. For example, of the five major classes of heparan sulfate proteoglycans, three are located in the ECM (perlecan, agrin, and type XVIII collagen) and two are cell-surface proteins. The latter include integral membrane proteins (syndecans) and GPI-anchored proteins (glypicans); the GAG chains in both types of cell-surface proteoglycans extend into the extracellular space. The sequences and lengths of proteoglycan core proteins vary considerably, and the number of attached GAG chains ranges from just a few to more than 100. Moreover, a core protein is often linked to two different types of GAG chains, generating a “hybrid” proteoglycan. The basal laminal proteoglycan perlecan is primarily a heparan sulfate proteoglycan with three to four GAG chains, although it can sometimes have a bound chondroitin sulfate chain. Additional diversity in proteoglycans occurs because the numbers, compositions, and sequences of the GAG chains attached to otherwise identical core proteins can differ considerably. Laboratory generation and analysis of mutants with defects in proteoglycan production in Drosophila melanogaster, C. elegans, and mice have clearly shown that proteoglycans play critical roles in development; for example, as participants in various signaling pathways (see Chapter 16 for examples in the TGF-β and Wnt pathways).
Syndecans are cell-surface proteoglycans expressed by epithelial and nonepithelial cells that bind to collagens and multi-adhesive matrix proteins such as fibronectin, anchoring cells to the ECM. Like that of many integral membrane proteins, the cytosolic domain of syndecan interacts with the actin cytoskeleton and in some cases with intracellular regulatory molecules. In addition, cell-surface proteoglycans such as syndecan bind many protein growth factors and other external signaling molecules, thereby helping to regulate cellular metabolism and function. For instance, syndecans in the hypothalamic region of the brain modulate feeding behavior in response to food deprivation. They do so by participating in the binding to cell-surface receptors of antisatiety peptides that help control feeding behavior. In the fed state, the syndecan extracellular domain decorated with heparan sulfate GAG chains is released from the cell surface by proteolysis, thus suppressing the activity of the antisatiety peptides and feeding behavior. In mice engineered to overexpress the syndecan-1 gene in the hypothalamic region of the brain and other tissues, normal control of feeding by antisatiety peptides is disrupted, and the animals overeat and become obese.