Connections Between the ECM and Cytoskeleton Are Defective in Muscular Dystrophy
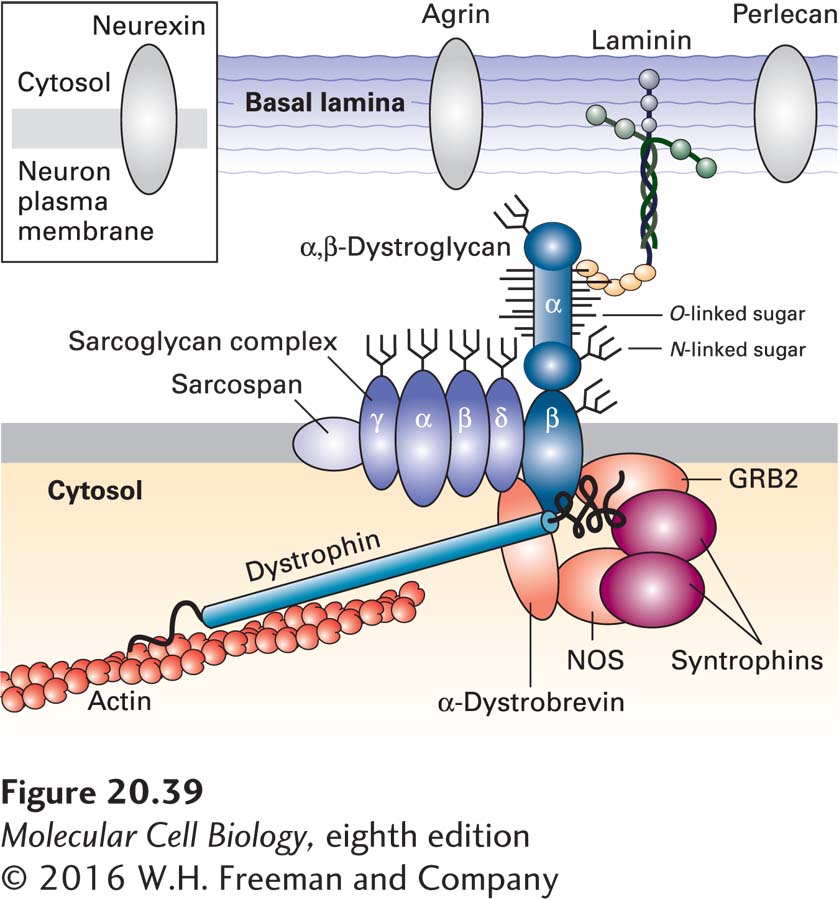
The importance of the adhesion receptor–mediated linkage between ECM components and the cytoskeleton is highlighted by a set of hereditary muscle-wasting diseases, collectively called muscular dystrophies. Duchenne muscular dystrophy (DMD), the most common type, is a sex-linked disorder, affecting 1 in 3300 boys, that results in cardiac or respiratory failure, usually in the late teens or early twenties. The first clue to understanding the molecular basis of this disease came from the discovery that people with DMD carry mutations in the gene encoding a protein named dystrophin. This very large protein was found to be a cytosolic adapter protein that binds to actin filaments and to an adhesion receptor called dystroglycan (Figure 20-39).
FIGURE 20-39The dystrophin glycoprotein complex (DGC) in skeletal muscle cells. This schematic model shows that the DGC comprises three subcomplexes: the α, β dystroglycan subcomplex; the sarcoglycan/sarcospan subcomplex of integral membrane proteins; and the cytosolic adapter subcomplex comprising dystrophin, other adapter proteins, and signaling molecules. Through its O-linked matriglycan sugars (see Figure 20-30c), α-dystroglycan binds to components of the basal lamina, such as laminin and perlecan, and to cell-surface proteins, such as neurexin in neurons. Dystrophin—the protein that is defective in Duchenne muscular dystrophy—links β-dystroglycan to the actin cytoskeleton, and α-dystrobrevin links dystrophin to the sarcoglycan/sarcospan subcomplex. Nitric oxide synthase (NOS) produces nitric oxide, a gaseous signaling molecule, and GRB2 is a component of signaling pathways activated by certain cell-surface receptors (see Chapter 15). See S. J. Winder, 2001, Trends Biochem. Sci.26:118; D. E. Michele and K. P. Campbell, 2003, J. Biol. Chem.278(18):15457–15460; and T. Yoshida-Moriguchi and K. P. Campbell, 2015, Glycobiology25:702–713.
Dystroglycan is synthesized as a large glycoprotein precursor that is proteolytically cleaved into two subunits soon after it is synthesized and before it moves to the cell surface. The α subunit is an extracellular peripheral membrane protein, and the β subunit is a transmembrane protein whose extracellular domain associates with the α subunit (see Figure 20-39). Multiple O-linked oligosaccharides are attached covalently to the side-chain hydroxyl groups of serine and threonine residues in the α subunit. Some of these linkages are unlike those in the most abundant O-linked oligosaccharides (also called mucin-like oligosaccharides), in which an N-acetylgalactosamine (GalNAc) is the first sugar in the chain linked directly to the hydroxyl group of the side chain of serine or threonine (see Figure 20-30b), or the linkage in proteoglycans (see Figure 20-30a). Instead, some of the more than 20 O-linked chains in dystroglycan are directly linked to the hydroxyl group via a mannose sugar (see Figure 20-30c). Some of these O-mannose-linked chains on dystroglycans have a phosphate group and six additional sugars attached to the mannose. A GAG-like polymer of xylose-glucuronic acid disaccharides called matriglycan is added at the end of this oligosaccharide. Matriglycan addition to dystroglycan in the Golgi complex, which is catalyzed by an enzyme called LARGE, requires the prior addition of the phosphorylated O-mannose-linked trisaccharide shown in Figure 20-30c.
Page 965
The O-linked matriglycan binds to various components of the basal lamina, including the LG domains of the multi-adhesive matrix protein laminin (see Figure 20-24) and the proteoglycans perlecan and agrin. The neurexins, a family of adhesion molecules expressed by neurons, are also bound via O-mannose-linked sugars.
The transmembrane segment of the dystroglycan β subunit associates with a complex of integral membrane proteins; its cytosolic domain binds dystrophin and other adapter proteins as well as various intracellular signaling proteins (see Figure 20-39). The resulting large, heteromeric assemblage, the dystrophin glycoprotein complex (DGC), links the ECM to the actin cytoskeleton and to signaling pathways within muscle and other types of cells. For instance, the signaling enzyme nitric oxide synthase (NOS) is associated through syntrophin with the DGC in skeletal muscle. The rise in intracellular Ca2+ during muscle contraction activates NOS to produce nitric oxide (NO), a signaling molecule that diffuses into smooth muscle cells surrounding nearby blood vessels. NO promotes smooth muscle relaxation, leading to a local rise in the flow of blood supplying nutrients and oxygen to the skeletal muscle. Heart (cardiac) muscle contraction may be influenced by similar NOS-syntrophin interactions.
Mutations in dystrophin, other DGC components, laminin, or the multiple enzymes that effect the addition of matriglycan to dystroglycan can all disrupt the DGC-mediated link between the exterior and the interior of muscle cells and cause muscular dystrophies. In addition, dystroglycan mutations have been shown to greatly reduce the clustering of acetylcholine receptors on muscle cells at the neuromuscular junctions, which is also dependent on the basal lamina proteins laminin and agrin. These and possibly other effects of DGC defects apparently lead to a cumulative loss of the mechanical stability of muscle cells as they undergo contraction and relaxation, resulting in deterioration of the cells and muscular dystrophy.
Dystroglycan provides an elegant—and medically relevant—example of the intricate networks of connectivity in cell biology. Dystroglycan was originally discovered in the context of studying muscular dystrophy. However, it was later shown to be expressed in nonmuscle cells and, through its binding to laminin, to play a key role in the assembly and stability of at least some basement membranes. Thus it is essential for normal development. Additional studies led to its identification as a cell-surface receptor for the virus that causes the frequently fatal human disease Lassa fever and other related viruses, all of which bind via matriglycan, the oligosaccharide on dystroglycan that mediates its binding to laminin. Furthermore, dystroglycan is the receptor on specialized cells in the nervous system—Schwann cells—to which binds the pathogenic bacterium Mycobacterium leprae, the organism that causes leprosy.