Leukocyte Movement into Tissues Is Orchestrated by a Precisely Timed Sequence of Adhesive Interactions
In adult organisms, several types of white blood cells (leukocytes) participate in defense against infection caused by bacteria and viruses and respond to tissue damage due to trauma or inflammation. To fight infection and clear away damaged tissue, these cells must move rapidly from the blood, where they circulate as unattached, relatively quiescent cells, into the underlying tissue at sites of infection, inflammation, or damage. We know a great deal about the movement into tissue, termed extravasation, of four types of leukocytes: neutrophils, which release several antibacterial proteins; monocytes, the precursors of macrophages, which can engulf and destroy foreign particles; and T and B lymphocytes, the antigen-recognizing cells of the immune system (see Chapter 23).
Extravasation requires the successive formation and breakage of cell-cell contacts between leukocytes in the blood and endothelial cells lining the vessels. Some of these contacts are mediated by selectins, a family of CAMs that mediate leukocyte–vascular endothelium interactions. Endothelial cells express P- and E-selectins on their blood-facing surfaces, activated platelets express P-selectin, and leukocytes express L-selectin. All selectins contain a Ca2+-dependent lectin domain, which is located at the distal end of the extracellular region of the molecule and recognizes particular sugars in glycoproteins or glycolipids (see Figure 20-2). For example, the primary ligand for P- and E-selectins is an oligosaccharide called the sialyl Lewis-x antigen, a part of longer oligosaccharides present in abundance on leukocyte glycoproteins and glycolipids.
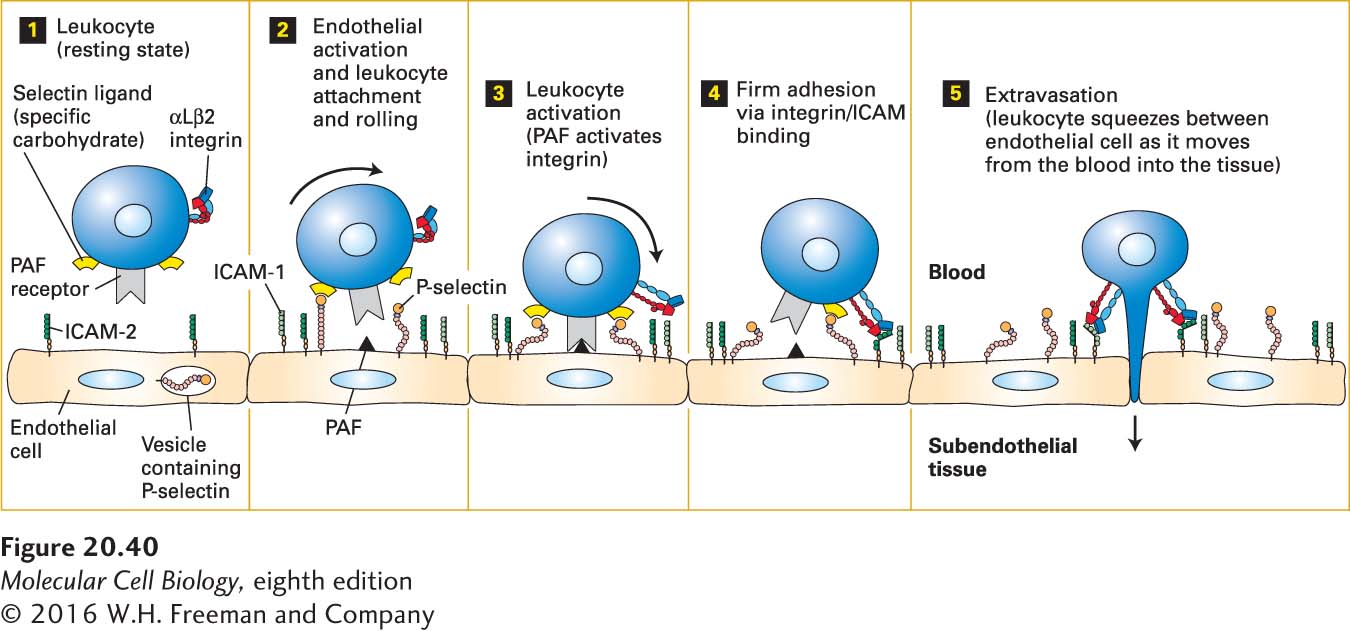
Figure 20-40 illustrates the basic sequence of cell-cell interactions leading to the extravasation of leukocytes. Various inflammatory signals released in areas of infection or inflammation first cause activation of the vascular endothelium. P-selectin exposed on the surfaces of activated endothelial cells mediates the weak adhesion of passing leukocytes. Because of the force of the blood flow and the rapid “on” and “off” rates of P-selectin binding to its ligands, these “trapped” leukocytes are slowed, but not stopped, and literally roll along the surface of the endothelium. Among the signals that promote activation of the endothelium are chemokines, a group of small secreted proteins (8–12 kDa) produced by a wide variety of cells, including endothelial cells and leukocytes.
FIGURE 20-40Endothelium-leukocyte interactions: activation, binding, rolling, and extravasation. Step 1: In the absence of inflammation or infection, leukocytes and endothelial cells lining blood vessels are in a resting state and not interacting. Step 2: Inflammatory signals released only in areas of inflammation, infection, or both activate resting endothelial cells, resulting in the movement of vesicle-sequestered selectins to the cell surface. The exposed selectins mediate weak binding of leukocytes by interacting with carbohydrate ligands on leukocytes. Blood flow forces the loosely bound leukocytes to roll along the endothelial surface of the blood vessel (curved arrow). Activation of the endothelium also causes synthesis of platelet-activating factor (PAF) and ICAM-1, both expressed on the endothelial cell surface. PAF and other, usually secreted, activators, including chemokines, then induce changes in the shapes of the leukocytes and activation of leukocyte integrins such as αLβ2, which is expressed by T lymphocytes (step 3). The subsequent tight binding between activated integrins on leukocytes and CAMs on the endothelium (e.g., ICAM-2 and ICAM-1) results in firm adhesion (step 4) and subsequent movement (extravasation) into the underlying tissue (step 5). See R. O. Hynes and A. Lander, 1992, Cell68:303.
For tight adhesion to occur between activated endothelial cells and leukocytes, β2-containing integrins on the surfaces of the leukocytes must be activated indirectly by chemokines or by other local activation signals such as platelet-activating factor (PAF). Platelet-activating factor is unusual in that it is a phospholipid rather than a protein; it is exposed on the surfaces of activated endothelial cells at the same time that P-selectin is exposed. The binding of PAF or other activators to their G protein–coupled receptors on leukocytes leads to activation of the leukocyte integrins (see Figure 20-38). These activated integrins then bind to distinct IgCAMs on the surfaces of endothelial cells. These IgCAMs include ICAM-2, which is expressed constitutively, and ICAM-1, whose synthesis is induced by activation. ICAM-1 does not usually contribute substantially to leukocyte adhesion to endothelial cells immediately after activation, but rather participates at later times in cases of chronic inflammation. The tight adhesion mediated by these Ca2+-independent integrin-ICAM interactions leads to the cessation of rolling and to the spreading of leukocytes on the surface of the endothelium; soon the adhered cells move between adjacent endothelial cells and into the underlying tissue. The extravasation step itself (also called transmigration or diapedesis; step 5 in Figure 20-40) requires the dissociation of otherwise stable adhesive interactions between endothelial cells that are primarily mediated by the CAM VE-cadherin. There is general agreement that the leukocyte interactions with endothelial cells mediated by CAMs initiate outside-in signaling in the endothelial cells that involves phosphorylation, activation of small GTPases, and an increase in cytosolic calcium concentration. These signals weaken or disrupt VE-cadherin-mediated inter-endothelial-cell adherens junctions and increase actin-myosin contraction, which pulls the endothelial cells apart, thus permitting the paracellular, amoeboid movement of the leukocyte between adjacent endothelial cells that is responsible for most extravasation.
Page 967
The selective adhesion of leukocytes to the endothelium near sites of infection or inflammation thus depends on the sequential appearance and activation of several different CAMs on the surfaces of the interacting cells. Different types of leukocytes express different integrins, though all contain the β2 subunit. Nonetheless, all leukocytes move into tissues by the general mechanism depicted in Figure 20-40.
Many of the CAMs used to direct leukocyte adhesion are shared among different types of leukocytes and target tissues. Yet often only a particular type of leukocyte is directed to a particular tissue. How is this specificity achieved? A three-step model has been proposed to account for the cell-type specificity of such leukocyte-endothelium interactions. First, endothelial activation promotes initial relatively weak, transient, and reversible binding (e.g., the interaction of selectins and their carbohydrate ligands). Without additional local activation signals, the leukocyte will quickly move on. Second, cells in the immediate vicinity of the site of infection or inflammation release or express chemical signals such as chemokines and PAFs that activate only special subsets of the transiently attached leukocytes, depending on the types of chemokine receptors those leukocytes express. Third, additional activation-dependent CAMs (e.g., integrins) engage their binding partners, leading to strong sustained adhesion. Only if the proper combination of CAMs, binding partners, and activation signals are engaged together with the appropriate timing at a specific site will a given leukocyte adhere strongly. Such combinatorial diversity and cross talk allows a small set of CAMs to serve diverse functions throughout the body—a good example of biological parsimony.
Leukocyte-adhesion deficiency is caused by a genetic defect in the synthesis of the integrin β2 subunit. People with this disorder are susceptible to repeated bacterial infections because their leukocytes cannot extravasate properly and thus cannot effectively fight infection within a tissue.
Some pathogenic viruses have evolved mechanisms to exploit cell-surface proteins that participate in the normal response to inflammation. For example, many of the RNA viruses that cause the common cold (rhinoviruses) bind to and enter cells through ICAM-1, and chemokine receptors can be important entry sites for human immunodeficiency virus (HIV), the cause of AIDS. Integrins appear to participate in the binding and/or internalization of a wide variety of viruses, including reoviruses (which cause fever and gastroenteritis, especially in infants), adenoviruses (which cause conjunctivitis and acute respiratory disease), and foot-and-mouth disease virus (which causes fever in cattle and pigs).