The Extracellular Matrix Participates in Adhesion, Signaling, and Other Functions
The extracellular matrix (ECM) is a complex combination of proteins and polysaccharides that is secreted and assembled by cells into a network in which the components bind to one another. The ECM is often involved in holding cells and tissues together. The composition, physical properties, and functions of the ECM are carefully controlled and can vary depending on the tissue type, its location, its physiological state, and chemical modifications of its components. These modifications include enzymatic phosphorylation, sulfation and desulfation, cross-linking, cleavage by proteases and glycosidases, and oxidation, as well as nonenzymatic addition of glucose (glycation).
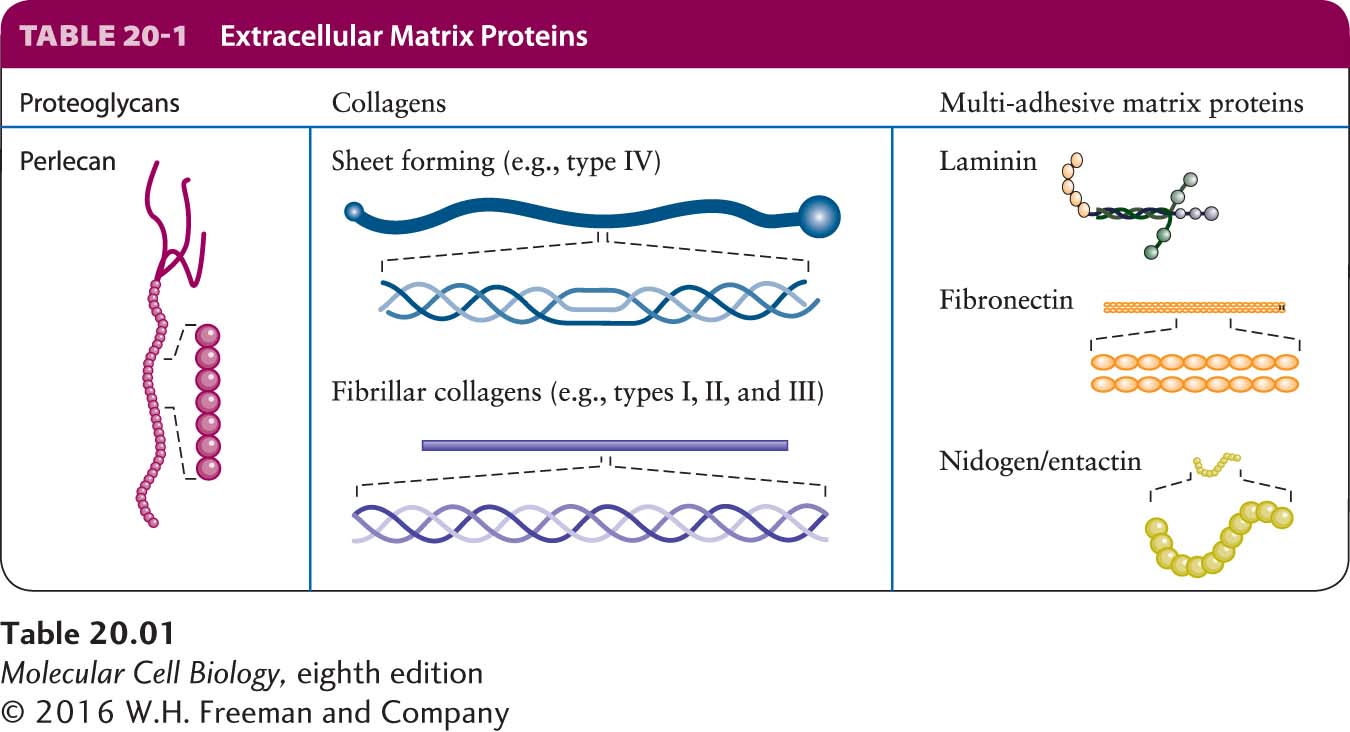
The ECM is usually sensed by cells as a consequence of binding to adhesion receptors on their plasma membranes, which then instruct the cells to behave appropriately in response to their environments or modulate the structure and function of the ECM based on the state of the cells. Different cells can bind to the same patch of ECM via their adhesion receptors and thus be indirectly bound together. ECM components include proteoglycans, a unique type of glycoprotein (a protein with covalently attached sugars); collagens and other proteins that often form fibers; soluble multi-adhesive matrix proteins; and others (Table 20-1). Multi-adhesive matrix proteins, such as fibronectin and laminin, are long, flexible molecules that contain multiple domains. They are responsible for binding various types of collagen, other matrix proteins, polysaccharides, and extracellular signaling molecules as well as adhesion receptors. These proteins are important organizers of the extracellular matrix. Through their interactions with adhesion receptors, they also regulate cell-matrix adhesion—and thus cell shape and behavior.
Cells contribute to the assembly of the ECM not only by secreting its components, but also by participating directly in the assembly of those components into complex structures containing large fibrils and amorphous macromolecules. Once assembled, the ECM often is not static, but rather highly dynamic in that its chemical, physical, and biological properties can be altered quantitatively or qualitatively as a consequence of cells secreting enzymes, such as proteases, and other molecules into the extracellular space. These alterations in the ECM, which are usually referred to as “remodeling,” can involve covalent chemical modifications (including chemical cross-linking of ECM molecules), partial or essentially complete proteolytic cleavage of ECM components, and addition of newly synthesized ECM molecules.
Page 926
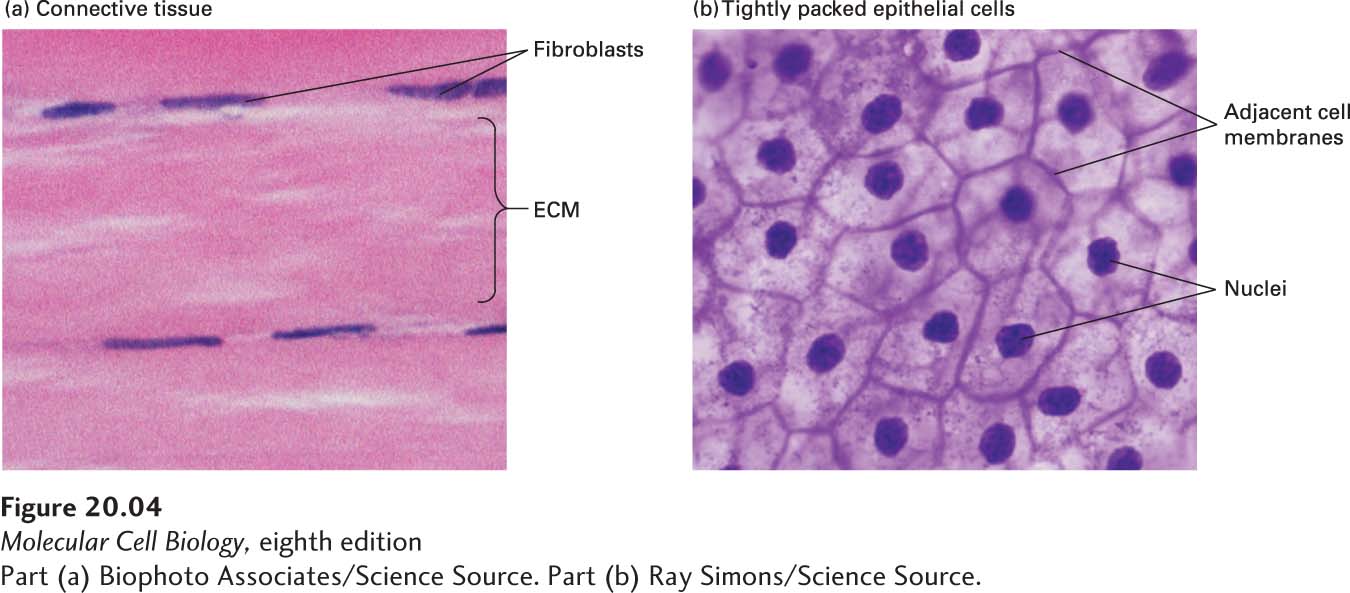
The relative volumes occupied by cells and their surrounding matrix vary greatly among different animal tissues. Some connective tissue, for instance, is mostly matrix with relatively few cells, whereas many other tissues, such as epithelia, are composed of very densely packed cells with relatively little matrix (Figure 20-4). The density of packing of the molecules within the ECM itself can also vary greatly.
[Part (a) Biophoto Associates/Science Source. Part (b) Ray Simons/Science Source.]
FIGURE 20-4Variation in the relative density of cells and ECM in different tissues. (a) Dense connective tissue contains mostly extracellular matrix consisting of tightly packed ECM fibers (pink) interspersed with rows of relatively sparse fibroblasts, the cells that synthesized this ECM (purple). (b) Squamous epithelium viewed from the top, showing epithelial cells tightly packed into a quilt-like pattern with the plasma membranes of adjacent cells close to one another and little ECM between the cells (see also Figure 20-10b).
[Part (a) Biophoto Associates/Science Source. Part (b) Ray Simons/Science Source.]
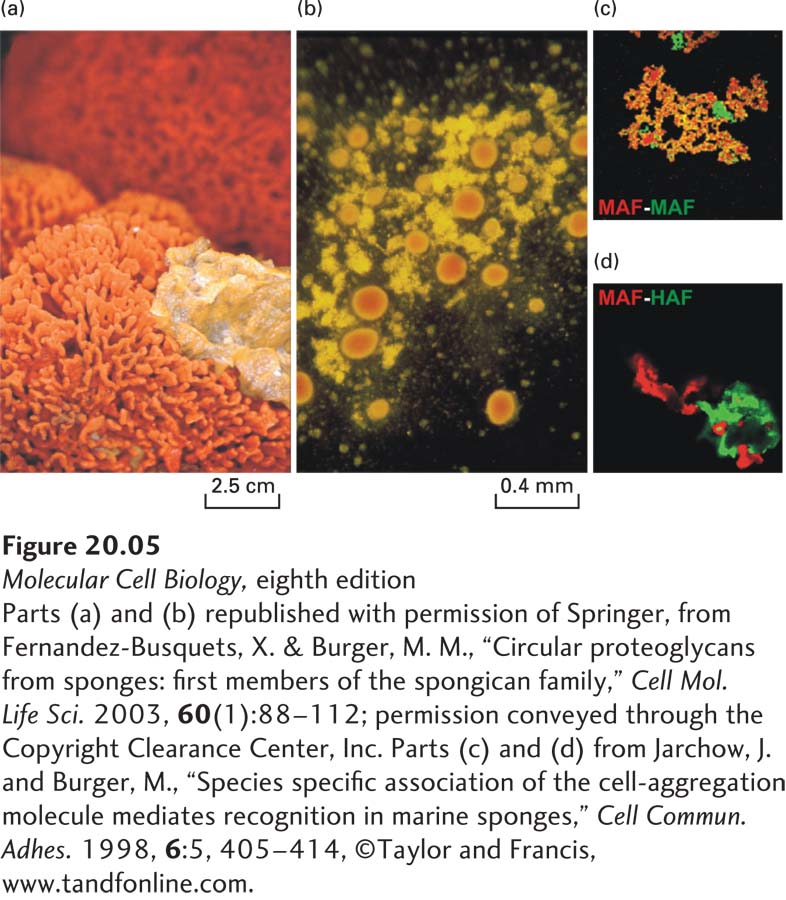
H. V. Wilson’s classic studies of adhesion in marine sponge cells showed conclusively that one primary function of the ECM is to literally hold tissue together. Figures 20-5a and 20-5b, which re-create Wilson’s classic work, show that when sponges are mechanically dissociated and individual cells from two sponge species are mixed, the cells of one species will adhere to one another, but not to cells from the other species. This specificity is due in part to species-specific adhesive proteins in the ECM that bind to the cells via adhesion receptors. These adhesive proteins can be purified and used to coat colored beads, which, when mixed, aggregate with one another with a specificity similar to that of intact sponge cells (Figure 20-5c, d).
EXPERIMENTAL FIGURE 20-5Mechanically separated marine sponges reassemble through species-specific homotypic cell adhesion. (a) Two intact sponges, Microciona prolifera (orange) and Halichondria panicea (yellow), growing in the wild. (b) After mechanical disruption and mixing of the individual cells from the two sponge species, their individual cells were allowed to reassociate for about 30 minutes with gentle stirring. The cells aggregated with species-specific homotypic adhesion, forming clumps of M. prolifera cells (orange) and H. panicea cells (yellow). (c) and (d) Red or green fluorescently labeled beads were coated with the proteoglycan aggregation factor (AF) from the ECM of either M. prolifera (MAF) or H. panicea (HAF). (c) When beads of both colors were coated with MAF, they all aggregated together, forming yellow aggregates (combination of red and green). (d) MAF (red) and HAF (green) coated beads do not readily form mixed aggregates, but rather assemble into distinct clumps held together by homotypic adhesion. (Magnification 40×.)
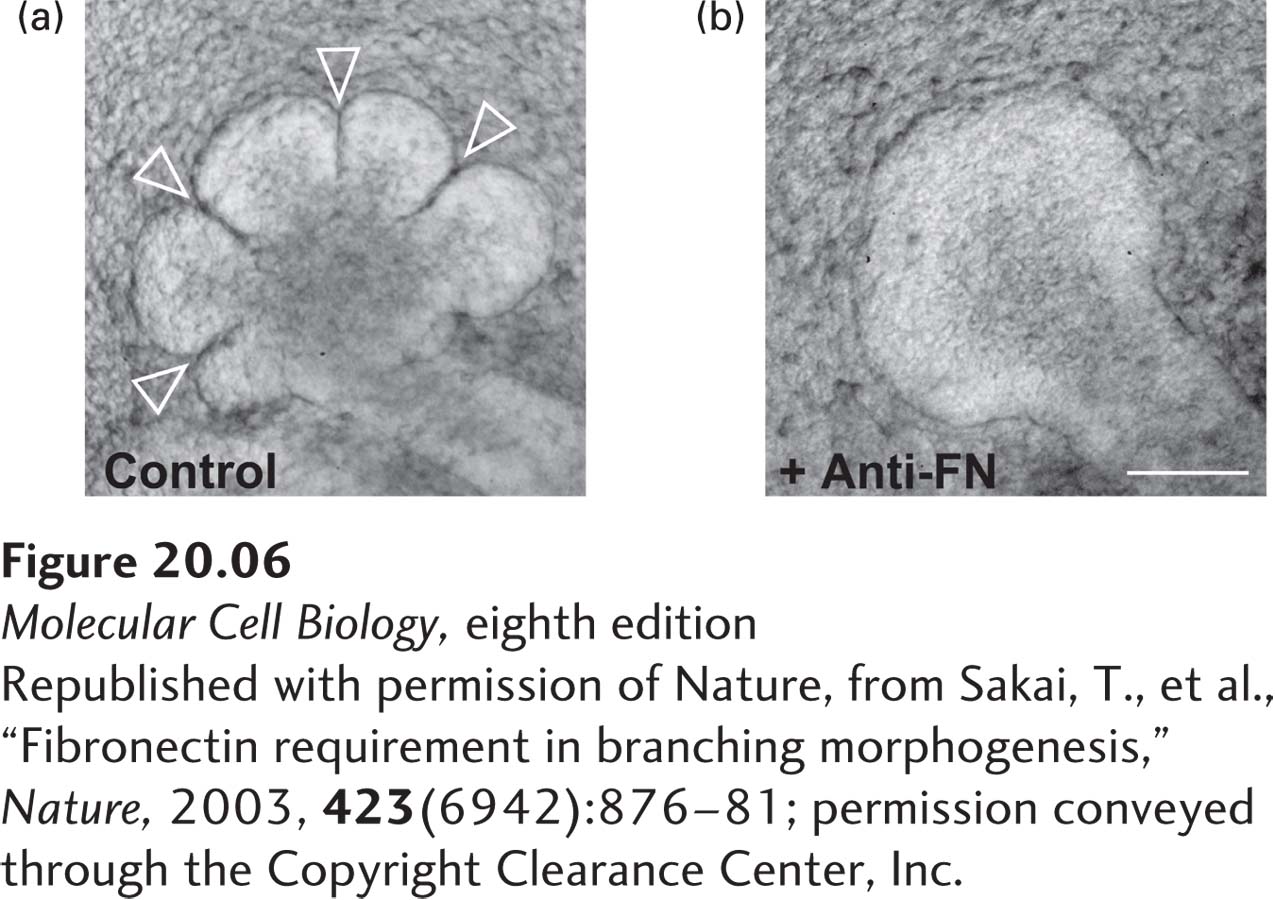
The ECM plays a multitude of other roles in addition to facilitating cell adhesion (Table 20-2). Different combinations of components tailor the ECM for specific purposes at different anatomic sites: strength in a tendon, strength and rigidity in teeth and bones, cushioning in cartilage, and transparency in the vitreous humor in the eyeball. The composition of the ECM also provides positional and signaling information for cells, letting a cell know where it is and what it should do. ECM remodeling can modulate the interactions of a cell with its environment. Furthermore, the ECM serves as a reservoir for many extracellular signaling molecules that control cell growth and differentiation. In addition, it provides a lattice through or on which cells either can move or are prevented from moving, particularly in the early stages of tissue assembly. Morphogenesis—the stage of embryonic development in which tissues, organs, and body parts are formed by cell movements and rearrangements—is critically dependent on cell-matrix adhesion as well as cell-cell adhesion. For example, cell-matrix interactions are required for branching morphogenesis (formation of branching structures) to form blood vessels, the air sacs in the lung, mammary and salivary glands, and other structures (Figure 20-6).
[Republished with permission of Nature, from Sakai, T., et al., “Fibronectin requirement in branching morphogenesis,” Nature, 2003, 423(6942):876–81; permission conveyed through the Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 20-6Antibodies to fibronectin block branching morphogenesis in developing mouse tissues. Immature salivary glands were isolated from murine embryos and allowed to undergo branching morphogenesis in vitro for 10 hours in the absence (a) or presence (b) of an antibody that binds to and blocks the activity of the ECM molecule fibronectin. Anti-fibronectin antibody (Anti-FN) treatment blocked branch formation (arrowheads). Inhibition of fibronectin’s adhesion receptor (an integrin) also blocks branch formation (not shown). Scale bar, 100 µm.
[Republished with permission of Nature, from Sakai, T., et al., “Fibronectin requirement in branching morphogenesis,” Nature, 2003, 423(6942):876–81; permission conveyed through the Copyright Clearance Center, Inc.]
Page 927
Page 928
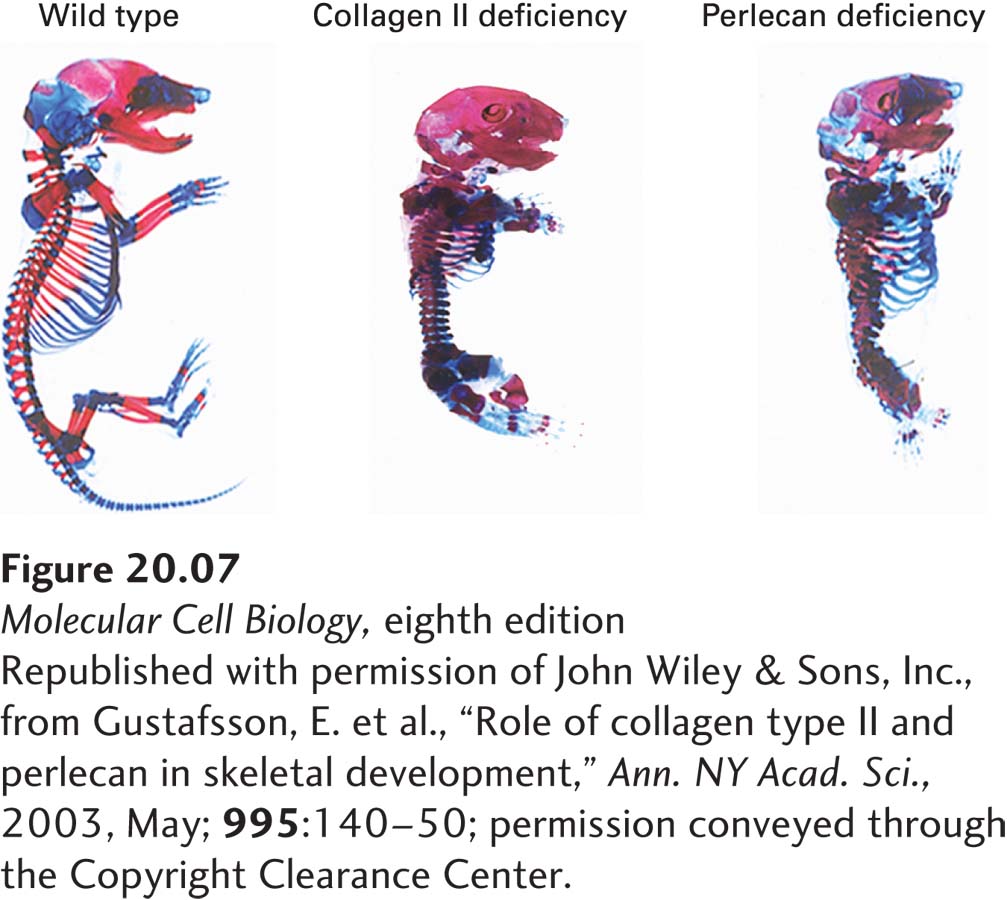
Disruptions in cell-matrix and cell-cell interactions can have devastating consequences for the development of tissues. Figure 20-7 shows the dramatic changes in the skeletal system of embryonic mice when the genes for either of two key ECM molecules, collagen II and perlecan, are inactivated. Disruptions in adhesion and ECM functions are also characteristic of various pathologies, including cardiovascular, musculoskeletal, kidney, skin, eye, and bone diseases as well as metastatic cancer, in which cancer cells leave their normal locations and spread throughout the body.
[Republished with permission of John Wiley & Sons, Inc., from Gustafsson, E. et al., “Role of collagen type II and perlecan in skeletal development,” Ann. NY Acad. Sci., 2003, May; 995:140–50; permission conveyed through the Copyright Clearance Center.]
EXPERIMENTAL FIGURE 20-7Inactivating the genes for some ECM proteins results in defective skeletal development in mice. These photographs show skeletons of normal (left), collagen II–deficient (center), and perlecan-deficient (right) murine embryos that were isolated and stained to visualize the cartilage (blue) and bone (red). Absence of these key ECM components leads to dwarfism, with many skeletal elements shortened and disfigured.
[Republished with permission of John Wiley & Sons, Inc., from Gustafsson, E. et al., “Role of collagen type II and perlecan in skeletal development,” Ann. NY Acad. Sci., 2003, May; 995:140–50; permission conveyed through the Copyright Clearance Center.]
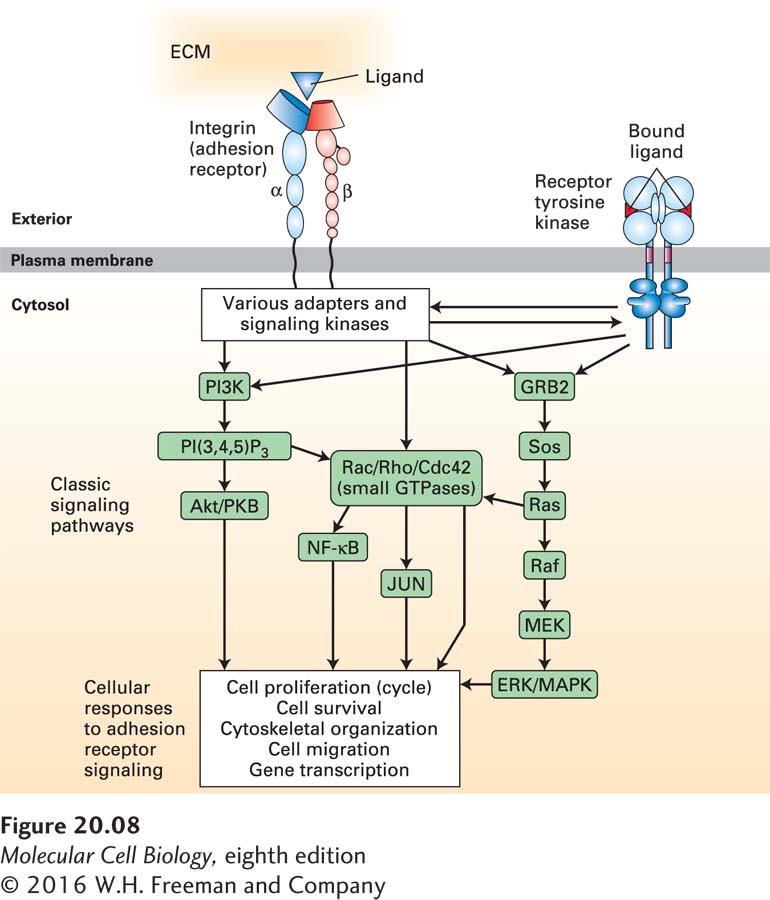
Although many CAMs and adhesion receptors were initially identified and characterized because of their adhesive properties, they also play major roles in signaling, using many of the pathways discussed in Chapters 15 and 16. Figure 20-8 illustrates how one adhesion receptor, integrin, physically and functionally interacts, via adapters and signaling molecules, with a broad array of intracellular signaling pathways to influence cell survival, gene transcription, cytoskeletal organization, cell motility, and cell proliferation. Conversely, changes in the activities of signaling pathways inside cells can influence the structures of CAMs and adhesion receptors—for example, by altering adapter binding to the cytosolic portions of the CAMs—and so modulate their ability to interact with other cells and with the ECM. Thus outside-in and inside-out signaling involve numerous interconnected pathways.
FIGURE 20-8Integrin adhesion receptor–mediated signaling pathways control diverse cell functions. Binding of integrins to their ligands induces conformational changes in their cytoplasmic domains, directly or indirectly altering their interactions with cytoplasmic proteins (outside-in signaling). These cytoplasmic proteins include adapter proteins (e.g., talins, kindlins, paxillin, vinculin) and signaling kinases [Src-family kinases, focal adhesion kinase (FAK), integrin-linked kinase (ILK)] that transmit signals via diverse signaling pathways, thereby influencing cell proliferation, cell survival, cytoskeletal organization, cell migration, and gene transcription. Components of several signaling pathways, some of which are associated directly with the plasma membrane, are shown in green boxes. Many of the components of the pathways shown here are shared with other cell-surface-activated signaling pathways (e.g., receptor tyrosine kinases shown on the right) and are discussed in Chapters 15 and 16. In turn, intracellular signaling pathways can, via adapter proteins, modify the ability of integrins to bind to their extracellular ligands (inside-out signaling). See W. Guo and F. G. Giancotti, 2004, Nat. Rev. Mol. Cell Biol.5:816–826, and R. O. Hynes, 2002, Cell110:673–687.