Polarized Membrane Traffic Allows Yeast to Grow Asymmetrically During Mating
One of the simplest and best-studied forms of cell asymmetry occurs when budding yeast cells mate. As we have seen, yeast can exist in a haploid state (with a single copy of each chromosome) or a diploid state (with two copies of each chromosome). The haploid state can exist in either of two mating types (“sexes”), a or α. The preferred state of yeast in nature is the diploid state, so a cells are always looking to mate with α cells to restore the diploid state (see Figure 1-23). Each mating type secretes a specific mating pheromone—a cells secrete a factor and α cells secrete α factor—and each expresses on its surface a receptor that senses the pheromone of the opposite mating type. Thus a cells have a receptor for α factor and α cells have a receptor for a factor. When cells of opposite mating types are placed near each other, the receptors on each cell bind and detect the pheromone cue of the other cell and determine its spatially highest concentration in order to know in which direction to mate. When the cells detect the opposite mating factor, two important processes occur. First, they synchronize their cell cycles by arresting at G0 so that when they mate, the two haploid genomes will be at the same stage of the cell cycle. Second, they target cell growth in the direction of the pheromone to assemble a mating projection called a shmoo. If shmooing cells of opposite mating types touch, they fuse at the shmoo tips, and the haploid nuclei come together to restore the diploid state.
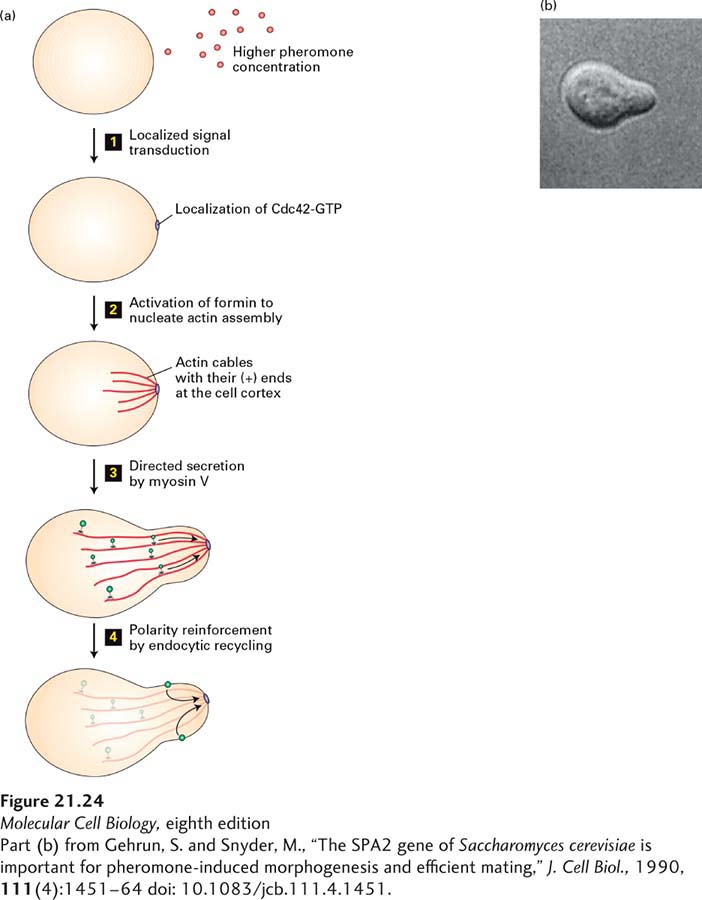
By looking for mutants in yeast haploids that cannot shmoo in response to the opposite mating pheromone, researchers have discovered how the asymmetric growth necessary for shmoo formation occurs (Figure 21-24). As might be anticipated, this mechanism initially involves a signal transduction pathway that establishes a polarized cytoskeleton, which in turn guides membrane traffic to the appropriate location for asymmetric growth. Activation of the mating-factor receptor—a typical G protein–coupled receptor (see Figure 15-12)—results in activation of the intrinsic polarity program, which in turn results in the localized accumulation and activation of Cdc42 in the region of the cell cortex closest to the pheromone source (Figure 21-24, step 1). This active Cdc42·GTP leads to the local activation of a formin protein (step 2). As we saw in Chapter 17, formin proteins nucleate the assembly of polarized actin filaments, whose (+) ends remain bound to the formin (see Figure 17-13). This process provides the tracks for the transport of secretory vesicles by a myosin V motor to the (+) ends of the filaments for localized growth and hence shmoo formation (step 3). Notice that this mechanism requires polarity proteins, which include Cdc42·GTP, to remain concentrated at the growing shmoo tip. To ensure that polarity is maintained during shmoo growth, a directed endocytic cycle is believed to exist. In this cycle, Cdc42 that has diffused away from the site of concentration may be internalized by endocytosis and transported back to the shmoo tip, thereby reinforcing polarity (step 4).
[Part (b) from Gehrun, S. and Snyder, M., “The SPA2 gene of Saccharomyces cerevisiae is important for pheromone-induced morphogenesis and efficient mating,” J. Cell Biol., 1990, 111(4):1451–64 doi: 10.1083/jcb.111.4.1451.]
FIGURE 21-24Mechanism of shmoo formation in yeast. (a) The haploid yeast cell must grow toward the highest concentration of mating factor of the opposite mating type, so it has a receptor on its surface that signals the location of the highest concentration. This signal induces the localization and activation of Cdc42 to generate a higher concentration of Cdc42·GTP at this site (step 1). The Cdc42·GTP locally activates a formin, which nucleates and elongates actin filaments from this site (step 2). Because formins bind to the (+) ends of actin filaments, the (+) ends are oriented toward Cdc42·GTP and thus the highest concentration of the mating factor. A myosin V motor transports secretory vesicles along the actin filaments, resulting in the growth of the shmoo (step 3). The polarity of the shmoo is reinforced by an endocytic cycle that constantly returns diffusing polarity factors, such as Cdc42, back along the actin filaments to the signal site (step 4). (b) DIC light-microscope image of a shmooing yeast cell.
[Part (b) from Gehrun, S. and Snyder, M., “The SPA2 gene of Saccharomyces cerevisiae is important for pheromone-induced morphogenesis and efficient mating,” J. Cell Biol., 1990, 111(4):1451–64 doi: 10.1083/jcb.111.4.1451.]