The Par Proteins Direct Cell Asymmetry in the Nematode Embryo
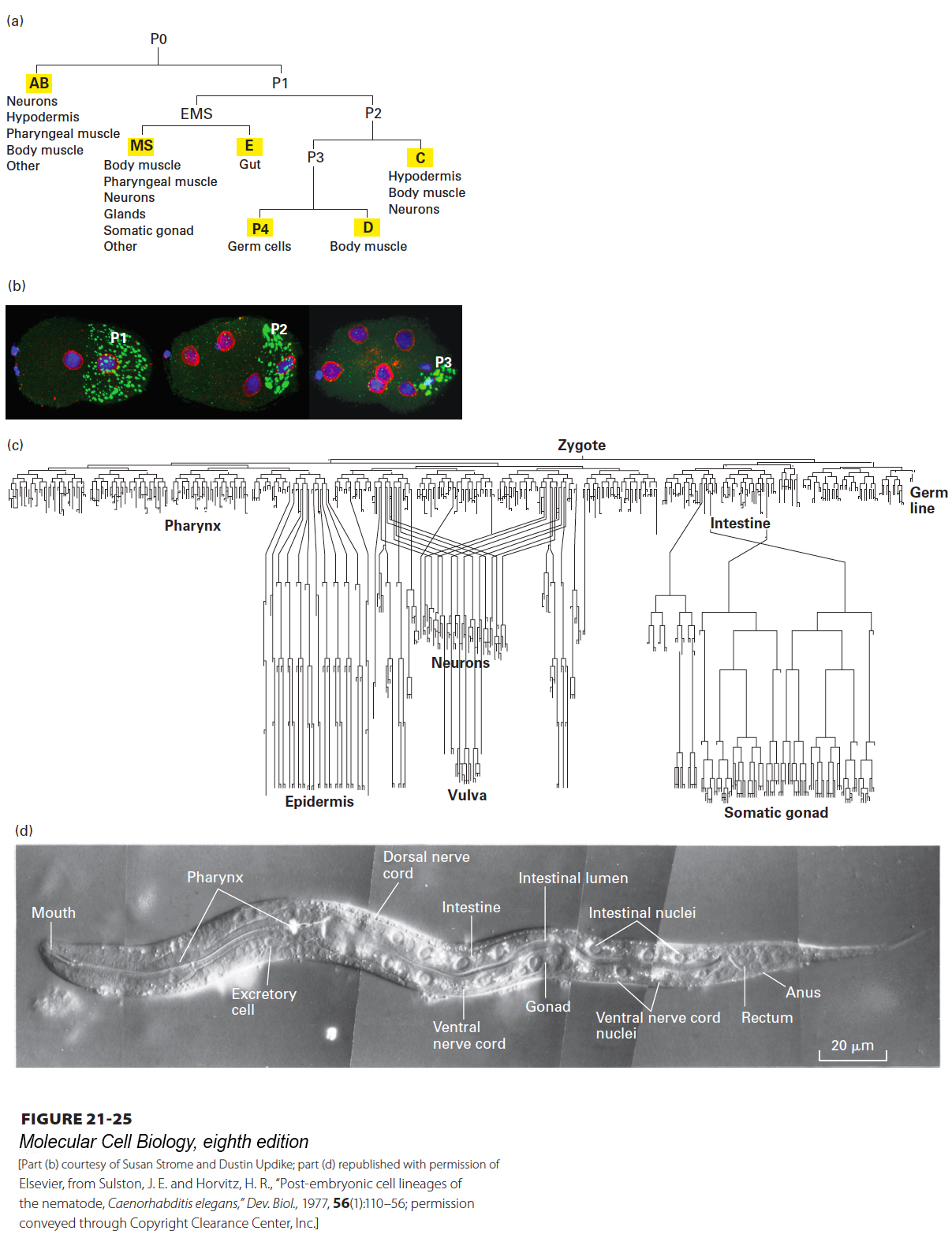
The nematode worm Caenorhabditis elegans has provided a powerful model system for understanding cell fate decisions. It was selected for study because the animal is transparent and has a rapid life cycle; it is easy to generate and characterize mutants; and the lineage of cells from the one-cell embryo to the adult is invariant (Figure 21-25a, c, d). A critical aspect of this lineage is the first cell division, in which the P0 cell—the fertilized egg, or zygote—gives rise to the AB and P1 cells by an asymmetric cell division; each of these two cells then gives rise to different lineages. Much is known about this first asymmetric division, which is where we focus our attention.
[Part (b) courtesy of Susan Strome and Dustin Updike; part (d) republished with permission of Elsevier, from Sulston, J. E. and Horvitz, H. R., “Post-embryonic cell lineages of the nematode, Caenorhabditis elegans,” Dev. Biol., 1977, 56(1):110–56; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 21-25Cell lineage in the nematode worm C. elegans. (a) Pattern of the first few divisions, starting with P0 (the zygote) and leading to formation of the six founder cells (yellow highlights). The first division is asymmetric, giving rise to the AB and P1 cells. The EMS cell is so named because it gives rise to most of the endoderm and mesoderm. The P4 lineage gives rise to the cells of the germ line. (b) Micrographs of two-, four-, and eight-cell embryos with DNA stained blue, the nuclear envelope red, and P granules green. The P1, P2, and P3 cells, which will give rise to the germ line, are indicated. (c) The full lineage of the entire body of the worm, showing some of the tissues formed. In this diagram, cell division is indicated by the splitting of a line, and the time of cell division is indicated in the vertical direction. (d) Newly hatched larva. Some of the 959 somatic-cell nuclei found in the adult hermaphrodite form can be seen in this micrograph obtained by differential-interference-contrast (DIC) microscopy.
[Part (b) courtesy of Susan Strome and Dustin Updike; part (d) republished with permission of Elsevier, from Sulston, J. E. and Horvitz, H. R., “Post-embryonic cell lineages of the nematode, Caenorhabditis elegans,” Dev. Biol., 1977, 56(1):110–56; permission conveyed through Copyright Clearance Center, Inc.]
Page 1004
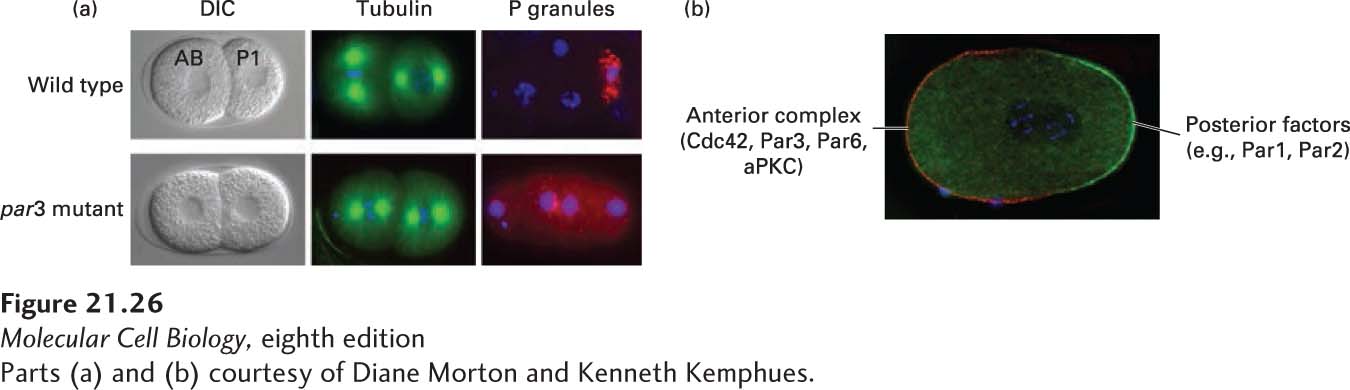
Before the first cell division, the zygote is visibly asymmetric: cytoplasmic complexes called P granules are concentrated at the cell end that will give rise to the posterior end of the embryo (Figure 21-25b). It turns out that during further cell divisions, these P granules always segregate to cells that will give rise to the germ line, where they ultimately play an important role in germ-line development. The first asymmetric division of the P0 cell gives rise to the P1 cell, containing the P granules, and the larger AB cell. Following that, at the two-cell stage, the mitotic spindles are arranged at right angles to one another so that the ensuing cell divisions are also at right angles to one another (Figure 21-26a). To begin to understand how this first essential asymmetric division occurs, mutations in six different genes were identified that resulted in a symmetric first division. Since the P granules were not partitioned correctly in these mutants, the genes identified in this study were called partition defective, or par, genes. In these mutants, P granules did not properly localize to the posterior end of the zygote, and the mitotic spindles were not oriented correctly in preparation for the second division (see Figure 21-26a). A key insight came when the products of the par genes were localized. In wild-type zygotes, many of the Par proteins localize either at the cortex of the anterior half of the cell or at the cortex of the posterior half. For example, Par3 (as part of a larger complex comprising Cdc42, Par3, Par6, and aPKC—atypical protein kinase C) localizes anteriorly, while Par2 and Par1 localize posteriorly (Figure 21-26b). Subsequent work has shown that mutually antagonistic interactions exist between these protein complexes; that is, if the Cdc42-Par3-Par6-aPKC complex is localized to one region, it excludes Par2, and vice versa. This is shown by the finding that the Par3-Par6-aPKC complex spreads over the whole cortex in par2 mutants and Par2 spreads over the whole cortex in par3 or par6 mutants. The molecular nature of this antagonism is not fully understood, but part of it is mediated by the protein kinase aPKC, which phosphorylates Par2 to inhibit its ability to bind to the anterior cortex.
[Parts (a) and (b) courtesy of Diane Morton and Kenneth Kemphues.]
EXPERIMENTAL FIGURE 21-26Par proteins are asymmetrically localized in the one-cell worm embryo. (a) DIC images of wild-type and par3 mutant embryos. Notice that in wild-type cells, the AB cell is larger than the P1 cell, whereas they are the same size in the par3 mutant. The par3 mutant also has a defect in spindle orientation (as seen by microtubule staining in green) and P-granule (red) segregation. DNA is stained blue. (b) Complementary localization of the anterior Par complex (Cdc42-Par3-Par6-aPKC) (red) and posterior determinants (green) in the one-cell embryo.
[Parts (a) and (b) courtesy of Diane Morton and Kenneth Kemphues.]
Page 1005
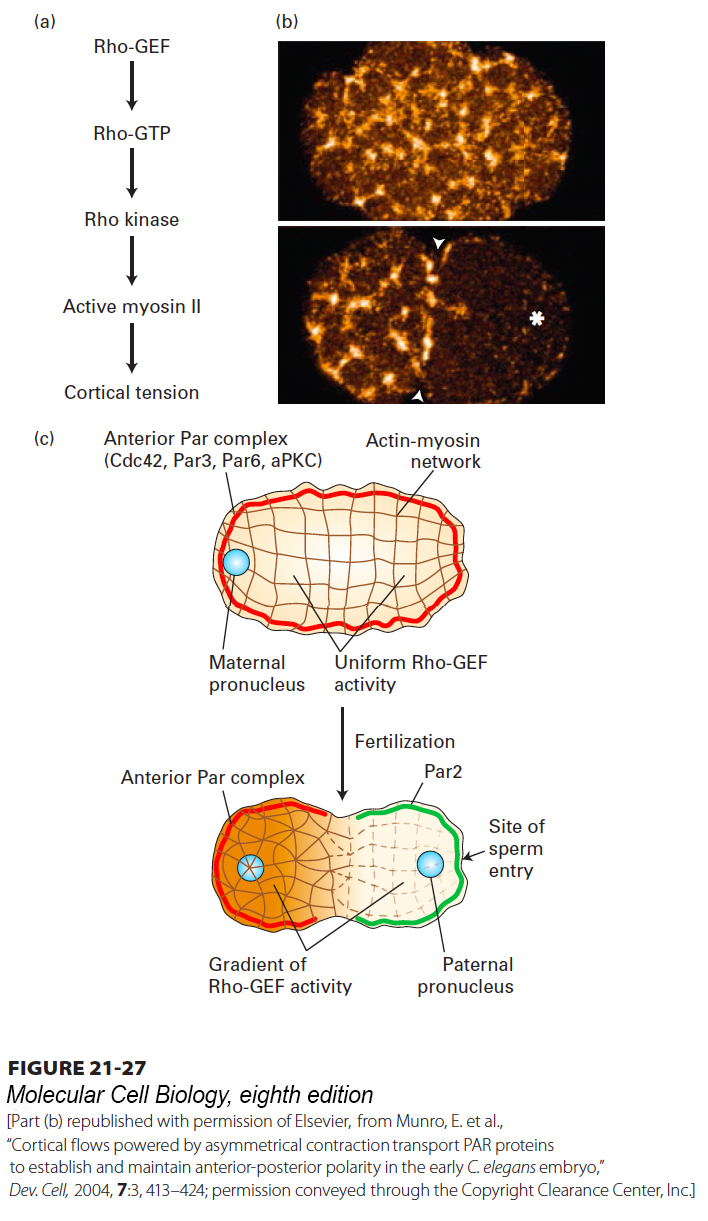
The unfertilized egg is symmetric, so what breaks this symmetry to generate a polarized zygote? It turns out that the position of the sperm after fertilization determines the posterior end. Prior to sperm entry, the entire egg cortex is under tension provided by an actin meshwork containing active myosin II. As we saw in Chapter 17, myosin II can form bipolar filaments that pull on actin filaments to generate tension, as is also seen in muscle and the contractile ring. Myosin II activity is regulated by a signal transduction pathway involving the small GTPase Rho (see Figure 17-43). In the unfertilized egg, Rho is maintained in its active Rho·GTP state by the uniform distribution of its activator, the guanine nucleotide exchange factor Rho-GEF. Rho·GTP activates Rho kinase, which phosphorylates the myosin light chain of myosin II to activate it (Figure 21-27a). Shortly after fertilization, an unknown signal from the sperm centrosome results in the local depletion of the Rho-GEF that is necessary to maintain active Rho. Thus the asymmetric position of the sperm centrosome defines the posterior region by depleting the Rho-GEF, thereby lowering the activity of myosin II. With this local reduction in contractile activity, the actin-myosin network contracts toward the anterior (Figure 21-27b), and as it does so, it drags (in an unknown manner) the anterior complex containing Par3, Par6, and aPKC to that end (Figure 21-27c). With the removal of the anterior complex, Par2 can now occupy the posterior cortex, and cell asymmetry is established.
[Part (b) republished with permission of Elsevier, from Munro, E. et al., “Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo,” Dev. Cell, 2004, 7:3, 413–424; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 21-27Mechanism of segregation of the anterior Par complex in the one-cell worm embryo. (a) Before fertilization, the cell cortex is under tension due to the activity of Rho-GEF, the guanine nucleotide exchange factor for the small GTPase Rho. Rho·GTP activates Rho kinase, which phosphorylates the regulatory light chain of myosin II to activate it. Together with actin filaments, the active myosin II maintains tension in the cell cortex. (b) Localization of myosin II before (top) and after (bottom) fertilization. The asterisk marks the region of sperm entry. (c) Before fertilization, Rho-GEF is uniformly active, the cortex is under tension from active myosin II, and the anterior Par complex (Cdc42-Par3-Par6-aPKC) is uniformly distributed around the cortex. Upon fertilization, Rho-GEF becomes locally reduced, resulting in local deactivation of myosin II. This deactivation generates unequal tension, so the actin–myosin network contracts toward the future anterior end, moving the anterior Par complex with it. Once the anterior complex is localized, factors such as Par2 associate with the posterior cell cortex. See D. St. Johnston and J. Ahringer, 2010, Cell141:757.
[Part (b) republished with permission of Elsevier, from Munro, E. et al., “Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo,” Dev. Cell, 2004, 7:3, 413–424; permission conveyed through the Copyright Clearance Center, Inc.]
Page 1006
It turns out that the master regulator Cdc42 is not needed for the initial asymmetry induced by actin-myosin network contraction. However, active Cdc42·GTP binds Par6 and is necessary for maintaining the anterior complex at the anterior end, although the mechanism for this localization is not yet clear. Recent work has also implicated an endocytic reinforcement cycle, as we discussed for yeast shmoo formation, to maintain polarity. Thus the steps of responding to a spatial cue, establishing asymmetry, and maintaining asymmetry are conserved features of both systems.