Formation of Synapses Requires Assembly of Presynaptic and Postsynaptic Structures
Axons extend from the cell body during development, guided by signals from other cells along the way so that the axon termini will reach the correct location (see Section 18.8). As axons grow, they come into contact with their potential target cells, such as dendrites of other neurons, and often at such sites synapses form. In the CNS, synapses with presynaptic specializations occur frequently all along an axon, and are called en passant (in passing) synapses; in contrast, motor neurons form synapses with muscle cells only at the axon termini.
Page 1049
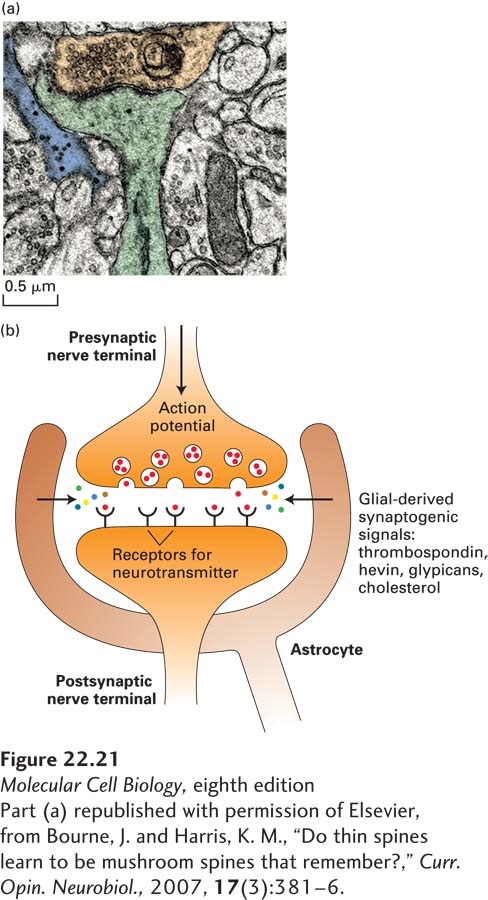
Neurons cultured in isolation will not form synapses very efficiently, but when glia are added, the rate of synapse formation increases substantially. Astrocytes and Schwann cells send protein signals to neurons to stimulate the formation of synapses and then help to preserve them. One such signal is thrombospondin (TSP), a component of the extracellular matrix; mice lacking two thrombospondin genes have only 70 percent of the normal number of synapses in their brains. Additional glial-derived signals are required for the formation of functional synapses, including glial-derived cholesterol, the extracellular matrix protein hevin, and glypicans (heparin sulfate proteoglycans). Mutual communication between neurons and the glia that surround them is frequent and complex, making the signals and information they carry an area of active research. New imaging approaches have revealed that astrocytes form a multitude of small branches that intercalate with neurons and ensheath synapses. Thus not only do astrocytes provide glial-derived factors to promote synapse formation, but many neuroscientists propose that the synapse, composed of presynaptic and postsynaptic partners, should be considered a tripartite synapse, composed not only of pre- and postsynaptic neuronal elements, but also of astrocytes (see Figure 22-21).
[Part (a) republished with permission of Elsevier, from Bourne, J. and Harris, K. M., “Do thin spines learn to be mushroom spines that remember?,” Curr. Opin. Neurobiol., 2007, 17(3):381–6.]
FIGURE 22-21Astrocytes and the tripartite synapse. (a) Many synapses are ensheathed by astroglial processes, as shown in this electron micrograph of a synapse in the rodent hippocampus. The postsynaptic compartment (dendrite and dendritic spine) is highlighted in green, the presynaptic terminal in orange, and astroglial process in blue. (b) Astrocytes not only ensheath the synapse, but they also secrete a number of factors that promote correct synapse formation. These include thrombospondin, hevin, glypicans, and cholesterol. Indicative of the importance of astrocytes in neuronal synapse formation, when neurons are grown in cell culture, they require astrocytes for proper synapse formation and development.
[Part (a) republished with permission of Elsevier, from Bourne, J. and Harris, K. M., “Do thin spines learn to be mushroom spines that remember?,” Curr. Opin. Neurobiol., 2007, 17(3):381–6.]
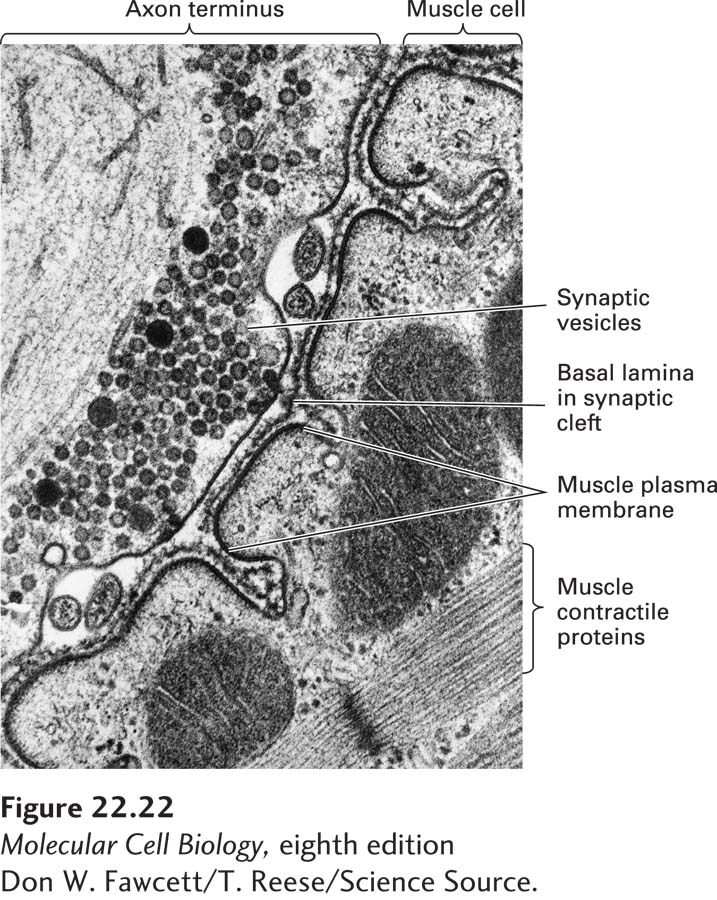
At the site of a synapse, the presynaptic neuron has hundreds to thousands of synaptic vesicles, some docked at the membrane and others waiting in reserve. The release of neurotransmitter into the synaptic cleft occurs in the active zone, a specialized region of the plasma membrane containing a remarkable assemblage of proteins whose functions include modifying the properties of the synaptic vesicles and bringing them into position for docking and fusing with the plasma membrane. Viewed by electron microscopy, the active zone has electron-dense material and fine cytoskeletal filaments (Figure 22-22). A similarly dense region of specialized structures is seen across the synapse in the postsynaptic cell, the postsynaptic density (PSD). Cell-adhesion molecules that connect pre- and postsynaptic cells keep the active zone and PSD aligned. After release of synaptic vesicles in response to an action potential, the presynaptic neuron retrieves synaptic vesicle membrane proteins by endocytosis both within and outside the active zone.
[Don W. Fawcett/T. Reese/Science Source.]
FIGURE 22-22Synaptic vesicles in the axon terminus near the region where neurotransmitter is released. In this longitudinal section through a neuromuscular junction, the basal lamina lies in the synaptic cleft separating the neuron from the muscle membrane, which is extensively folded. Acetylcholine receptors are concentrated in the postsynaptic muscle membrane at the top and part way down the sides of the folds in the membrane.
[Don W. Fawcett/T. Reese/Science Source.]
Page 1050
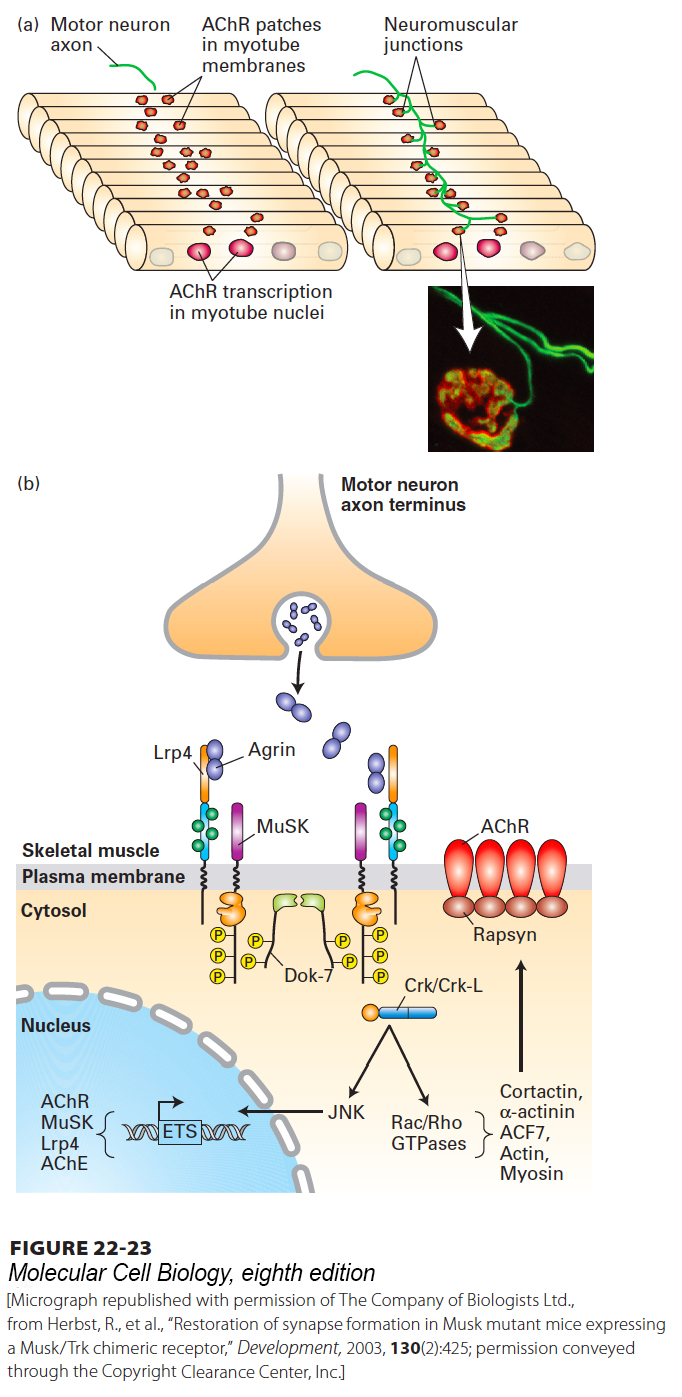
Synapse assembly has been extensively studied at the neuromuscular junction (NMJ) (Figure 22-23). At these synapses acetylcholine is the neurotransmitter produced by motor neurons, and its receptor, AChR, is produced by the postsynaptic muscle cell. Muscle cell precursors, myoblasts, put into culture will spontaneously fuse into multinucleate myotubes that look similar to normal muscle cells. As myotubes form, AChR is produced near the center of the cell and inserted into the myotube plasma membrane, forming diffuse membrane patches (Figure 22-23a).
[Micrograph republished with permission of The Company of Biologists Ltd., from Herbst, R., et al., “Restoration of synapse formation in Musk mutant mice expressing a Musk/Trk chimeric receptor,” Development, 2003, 130(2):425; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 22-23Formation of the neuromuscular junction. (a) Motor neuron-myotube interactions. Following fusion of myoblasts to form multinucleate myotubes, the nuclei synthesize acetylcholine receptor (AChR) mRNA. The nuclei near the center of each muscle fiber synthesize significantly more AChR mRNA than other nuclei. AChRs together with MuSK receptor kinases accumulate in membrane patches near the center of the cell, the prospective synaptic region of the muscle, prior to and independent of innervation; the cell is said to be “prepatterned.” The motor neuron axon termini grow toward these AChR clusters and secrete the glycoprotein Agrin. Agrin, in turn, induces clustering of the AChRs (dark red) and MuSKs around the axon termini (green), forming the neuromuscular junction. (Inset) Micrograph of a synapse from a postnatal (3-week-old) mouse, viewed by staining for axons (neurofilament) and synaptic vesicles (synaptophysin), shown together in green, and AChRs, shown in red. (b) Signaling downstream of Agrin receptors. Motor axons secrete Agrin, which stabilizes postsynaptic differentiation by binding LRP4 and activating MuSK kinase activity. Phosphorylation of tyrosines in the MuSK juxtamembrane region, indicated by yellow P in circle, stimulates recruitment and tyrosine phosphorylation of Dok-7, an adapter protein that is expressed selectively in muscle, which forms a dimer, stimulates MuSK kinase activity, and recruits the adapter protein Crk/Crk-L. Crk/Crk-L is essential to activate a Rac/Rho- and Rapsyn-dependent pathway for clustering AChRs opposite the presynaptic axon termini; this pathway involves several cytoskeletal proteins including actin and myosins. The pathway for synapse-specific transcription is less well understood but likely involves JNK kinase-dependent activation of ETS-family transcription factors that stimulate expression of multiple genes encoding synaptic proteins such as acetylcholine receptors, MuSK, LRP4, and acetylcholinesterase (AChE), the extracellular enzyme that localizes to the synaptic cleft and that degrades acetylcholine to choline and acetate.
[Micrograph republished with permission of The Company of Biologists Ltd., from Herbst, R., et al., “Restoration of synapse formation in Musk mutant mice expressing a Musk/Trk chimeric receptor,” Development, 2003, 130(2):425; permission conveyed through the Copyright Clearance Center, Inc.]
The formation of the neuromuscular synapse is a multistep process requiring signaling interactions between motor neurons and muscle fibers. A key player is MuSK, a receptor tyrosine kinase that is localized in the diffuse AChR-rich patches of the myotube plasma membrane. In ways that are not known, MuSK both induces clustering of AChRs and serves to attract the termini of growing motor neuron axons. For example, knockdown of MuSK inhibits both processes, while overexpression of MuSK in cultured muscle cells induces motor neuron growth throughout the muscle and formation of excess synapses.
Page 1051
Another key player is Agrin, a glycoprotein synthesized by developing motor neurons, transported in vesicles along axon microtubules, and secreted near the developing myotubes. Agrin binds to LRP4, a single-pass membrane-spanning protein; this stimulates an association between LRP4 and MuSK and increases MuSK kinase activity (Figure 22-23b). This leads to activation of several downstream signal transduction pathways, one of which leads to activation of Rac and Rho (see Section 17.3) and formation of clusters of AChRs with the cytoskeletal protein rapsyn; this interaction, together with binding of other cytoskeletal proteins including actin, leads to localization of AChRs opposite the nerve termini at the neuromuscular junction. The density of acetylcholine receptors in a mature synapse reaches ~10,000–20,000/µm2, while elsewhere in the plasma membrane the density is ~10/µm2. Another pathway, also not well understood, leads to activation of ETS-family transcription factors and stimulation of expression of multiple genes encoding synaptic proteins such as rapsyn and AChRs.
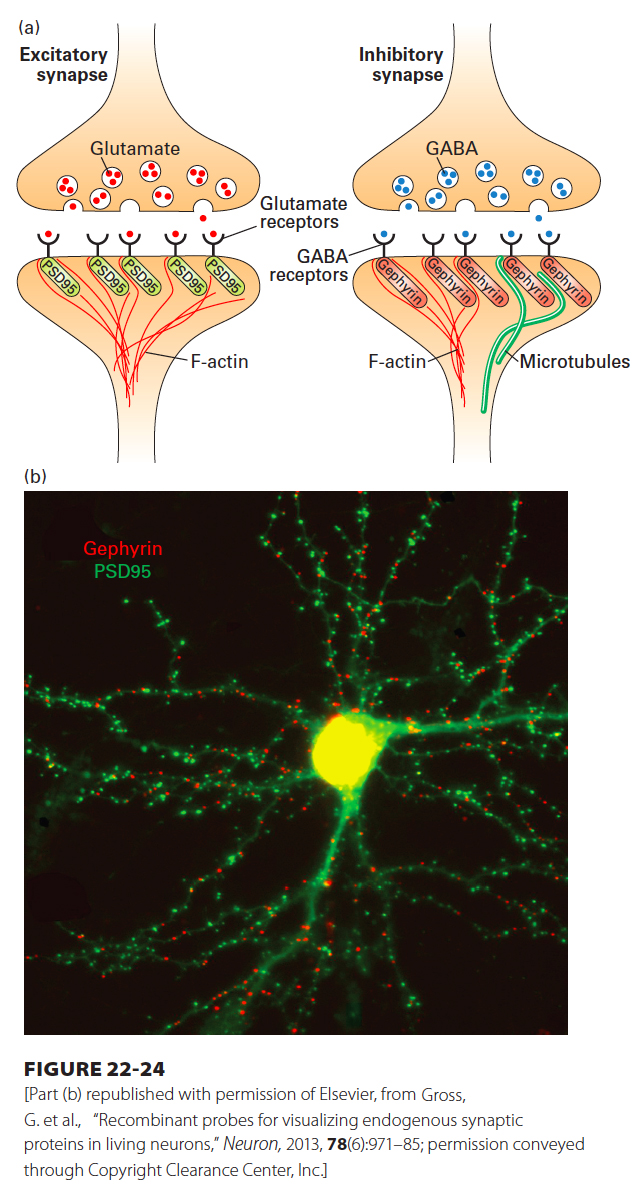
While the molecular mechanisms underlying the formation of synapses in the central nervous system are less well understood, the process appears to follow a similar logic, in which interactions between the pre- and postsynaptic compartments trigger a reorganization of already synthesized synaptic components. Analogous to the function of rapsyn in clustering ACh receptors at the neuromuscular synapse, distinct scaffolding proteins cluster neurotransmitter receptors at excitatory and inhibitory synapses in the central nervous system. A large PDZ-containing (see Chapter 20 for definition of PDZ domains) protein called PSD95 clusters glutamate receptors at excitatory synapses, while another scaffolding protein called gephryin clusters GABA and glycine receptors at inhibitory synapses (Figure 22-24). Pre- and postsynaptic compartments are linked by a network of trans-synaptic cell adhesion molecules, whose adhesive interactions are so strong that it is not possible to biochemically separate the presynaptic compartment from the postsynaptic compartment. These adhesion molecules include cadherins, immunoglobulin-containing cell adhesion molecules (described in Chapter 20), neurexins and neuroligins, ephrins, and Eph receptors. Synaptic adhesion molecules bind scaffolding proteins and cytoskeletal elements through their intracellular domains, promoting organization of protein complexes in both presynaptic and postsynaptic compartments. Mixed cell culture systems, in which primary neurons are cultured with non-neuronal cells expressing an adhesion molecule of interest, have been useful in demonstrating the ability of specific adhesion molecules to promote synapse assembly. For example, expression of the postsynaptic adhesion molecule neurexin in non-neuronal cells is sufficient to promote presynaptic specializations in axons, including formation of active zone components with clusters of synaptic vesicles.
[Part (b) republished with permission of Elsevier, from Gross, G. et al., “Recombinant probes for visualizing endogenous synaptic proteins in living neurons,” Neuron, 2013, 78(6):971–85; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 22-24PSD95 and gephyrin are scaffolding proteins for excitatory and inhibitory postsynaptic compartments, respectively. (a) The PDZ-containing protein PSD95 is part of the postsynaptic density at excitatory synapses and associates with glutamate receptors and with the actin cytoskeleton. The scaffolding protein gephyrin plays an analogous role at inhibitory synapses, where it associates with GABA receptors and with the microtubule and actin cytoskeletal networks. (b) PSD95 and gephyrin can be used to mark excitatory and inhibitory synapses, respectively. Shown in green are the PSD95-containing excitatory synapses and, in red, the gephyrin-containing inhibitory synapses that form on a single mouse cortical neuron in culture.
[Part (b) republished with permission of Elsevier, from Gross, G. et al., “Recombinant probes for visualizing endogenous synaptic proteins in living neurons,” Neuron, 2013, 78(6):971–85; permission conveyed through Copyright Clearance Center, Inc.]
Precise wiring in the nervous system involves not only synapse formation, but also synapse elimination. At birth, most animals are born with an excess number of synapses, and neural circuit maturation involves a critical process of synaptic pruning that eliminates a large number of synapses. Synaptic pruning is dependent on neural activity, and studies indicate that the weaker synapses are eliminated while the stronger synapses are maintained. For example, at birth each muscle cell is innervated by multiple motor neurons, but over time many of the motor neuron contacts are eliminated so that eventually each muscle cell is innervated by a single motor neuron. This process of synapse elimination is dependent on activity in the muscle cell: if the muscle cell is pharmacologically silenced, it never loses its polyneuronal innervation. The cell biological mechanisms underlying synapse elimination, including the mechanisms that specify which synapses should be eliminated and the process of elimination itself, are areas of active research. Many recent studies have uncovered roles for astrocytes and microglia in the phagocytosis of eliminated synapses during synaptic pruning.