Influx of Ca2+ Triggers Release of Neurotransmitters
The exocytosis of neurotransmitters from synaptic vesicles involves vesicle-targeting and fusion events similar to those that occur during the intracellular transport of secreted and plasma-membrane proteins (Chapter 14). However, two unique features critical to synapse function differ from other secretory pathways: (1) secretion is tightly coupled to arrival of an action potential at the axon terminus, and (2) synaptic vesicles are recycled locally to the axon terminus after fusion with the plasma membrane. Figure 22-26 shows the entire cycle whereby synaptic vesicles are filled with neurotransmitter, release their contents, and are recycled.
Depolarization of the plasma membrane cannot, by itself, cause synaptic vesicles to fuse with the plasma membrane. In order to trigger vesicle fusion, an action potential must be converted into a chemical signal—namely, a localized rise in the cytosolic Ca2+ concentration. The transducers of the electrical signals are voltage-gated Ca2+ channels localized to the region of the plasma membrane adjacent to the synaptic vesicles. The membrane depolarization due to arrival of an action potential opens these channels, permitting an influx of Ca2+ ions from the extracellular medium into the region of the axon terminus near the docked synaptic vesicles. Importantly, the rise in cytosolic Ca2+ is localized; it is also transient, as the excess Ca2+ is rapidly pumped out of the cell by plasma membrane Ca2+ pumps.
The bacterial voltage-dependent Na+ channel is likely the evolutionary ancestor of both voltage-dependent Na+ and Ca2+ channels. Specifically, the structures of the voltage-sensing module and of the pore module, as well the structural mechanisms underlying voltage-dependent activation and slow voltage-dependent inactivation, are conserved. The bacterial voltage-gated Ca2+ channel is unique among these channels in that it is selective for calcium ions. How does this channel achieve high and selective conductance for Ca2+ when the extracellular concentration of Na+ is 140 mM and the extracellular concentration of Ca2+ is only 2 mM? To determine the structural basis of this selectivity, scientists mutated residues in the selectivity pore of the bacterial voltage-dependent Na+ channel to residues that were found in voltage-gated Ca2+ channels. This turned the bacterial channel into a voltage-dependent Ca2+ channel whose structure could be solved by x-ray crystallography. These studies showed that mutation of a single serine residue in the selectivity filter to an aspartate converted the channel to one that was calcium selective, conducting calcium ions with a single shell of hydration. This mutation, together with additional mutations that changed the electronegativity of the pore, provided sufficient selectivity to conduct Ca2+ over Na+ despite the relative abundance of Na+ ions in the extracellular space. While the voltage-gated Na+ and K+ channels are the most important contributors to the generation of action potentials, the voltage-gated Ca2+ channels are essential for the conversion of electrical signals into chemical signals, since the influx of calcium into the neuron triggers a series of signal transduction cascades that leads to the release of synaptic vesicles and the transmission of the electrical signal from the one neuron to another neuron.
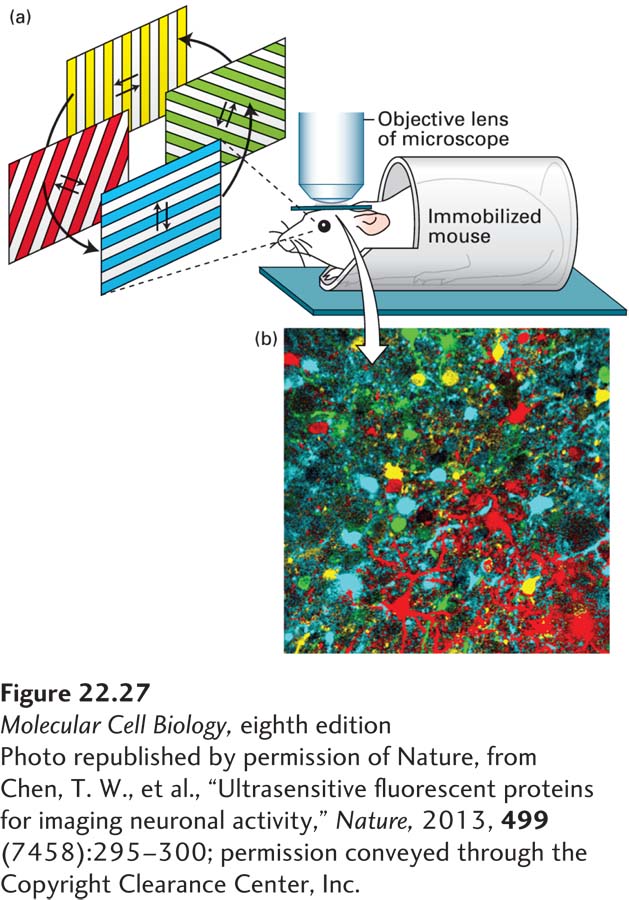
The development of fluorescent Ca2+ indicators has provided a powerful means of visualizing synaptic activity in neurons in culture and in intact neural circuits. As discussed in Chapter 4, these indicators are fluorescent molecules that change their fluorescence emission upon Ca2+ binding, and include both chemical indicators and genetically encoded indicators. Delivery or expression of Ca2+ indicators to neurons in a circuit allow experimenters to use time-lapse microscopy to monitor Ca2+ transients in hundreds of neurons and glia in real time. For example, expression of the genetically encoded Ca2+ indicator GCaMP6 in the visual cortex of mice, combined with presentation of visual stimuli and in vivo two-photon microscopy, has been used to identify the population of neurons that respond to specific orientations of visual information (Figure 22-27).
[Photo republished by permission of Nature, from Chen, T. W., et al., “Ultrasensitive fluorescent proteins for imaging neuronal activity,” Nature, 2013, 499(7458):295–300; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 22-27Calcium indicators allow visualization of activity in neural circuits. A genetically encoded calcium indicator was expressed in neurons in the mouse visual cortex. (a) A window was made in the skull of the mouse, and a microscope (indicated by the objective lens) was used to visualize calcium transients in populations of neurons in the visual cortex while the mouse was looking at gratings that moved in different directions. Individual neurons within the visual cortex respond to specific orientations of the gratings, as detected by elevations in calcium that are visualized as increases in the fluorescence of the calcium indicator. (b) Neurons were color coded according to the orientation that elicited increases in calcium (as shown below the photo). The neurons shown in yellow respond to horizontally moving gratings, and the neurons shown in cyan respond to vertically moving gratings, while the neurons shown in green and red respond to diagonally oriented gratings. This type of experiment reveals that individual neurons are tuned to specific orientations of visual stimuli.
[Photo republished by permission of Nature, from Chen, T. W., et al., “Ultrasensitive fluorescent proteins for imaging neuronal activity,” Nature, 2013, 499(7458):295–300; permission conveyed through the Copyright Clearance Center, Inc.]
Page 1055
A single action potential leads to exocytosis of about 10% of synaptic vesicles in a presynaptic terminal. Membrane proteins unique to synaptic vesicles then are specifically internalized by endocytosis, usually via the same types of clathrin-coated vesicles used to recover other plasma-membrane proteins by other types of cells. After the endocytosed vesicles lose their clathrin coat, they are rapidly refilled with neurotransmitter. The ability of many neurons to fire 50 times a second is clear evidence that the recycling of vesicle membrane proteins occurs quite rapidly. The machinery of endocytosis and exocytosis is highly conserved, and is described in more detail in Chapter 14.