Information Moves Along Axons as Pulses of Ion Flow Called Action Potentials
Nerve cells are members of a class of excitable cells, which also includes muscle cells, cells in the pancreas, and some others. Like all metazoan cells, excitable cells have an inside-negative voltage or electric potential gradient across their plasma membranes, the membrane potential (see Chapter 11). In excitable cells this potential can suddenly become zero or even reversed, with the inside of the cell positive with respect to the outside of the plasma membrane. The membrane voltage in a typical neuron, called the resting potential because it is the state when no signal is in transit, is established by Na+/ K+ ion pumps in the plasma membrane. These are the same ion pumps used by other cells to generate a resting potential. Na+/ K+ ion pumps use energy, in the form of ATP, to move positively charged Na+ ions out of the cell and K+ ions inward. Subsequent movement of K+ out of the cell through resting K+ channels results in a net negative charge inside the cell compared with the outside. The typical resting potential of a neuron is about −70 mV.
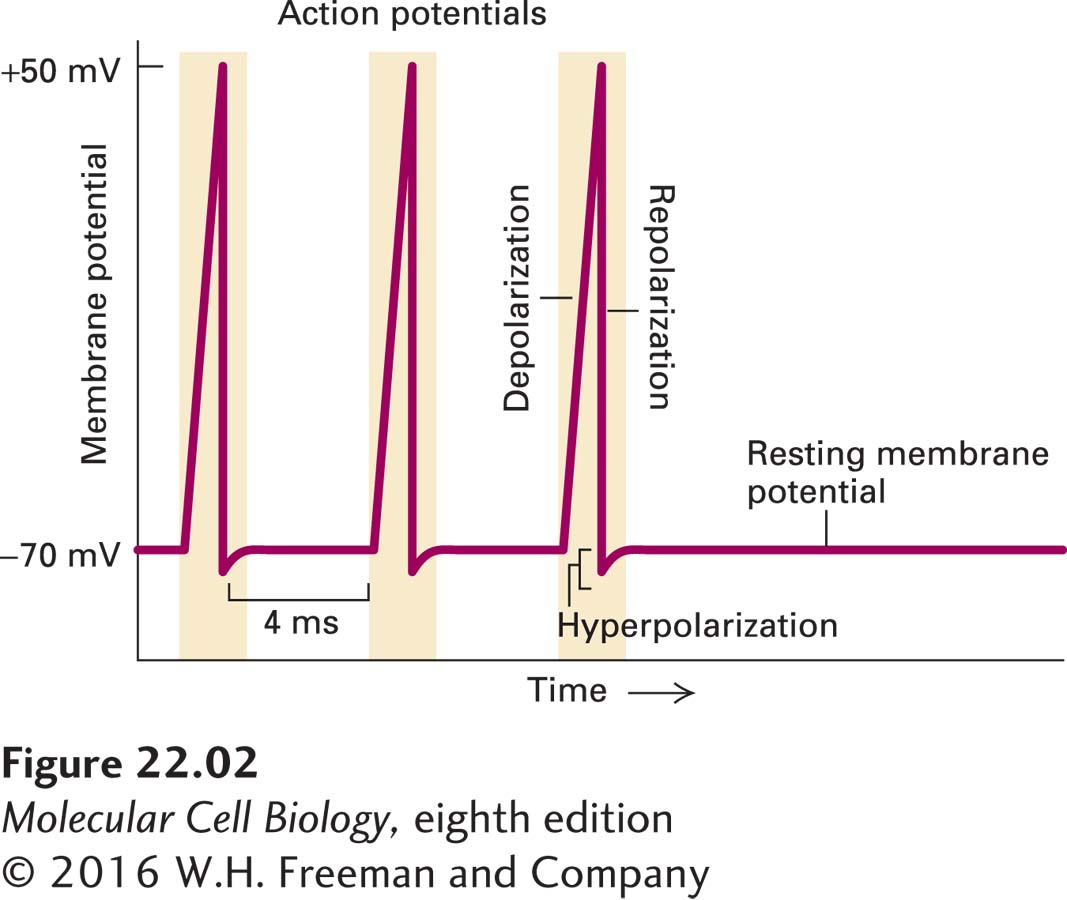
Neurons have a language all their own. They use their unique electrical properties to send signals. The signals take the form of brief local voltage changes, from inside-negative to inside-positive, an event designated depolarization. A powerful surge of depolarizing voltage change, moving from one end of the neuron to the other, is called an action potential. “Depolarization” is somewhat of a misnomer, since the neuron suddenly goes from inside-negative to neutral to inside-positive, which could be more accurately described as depolarization followed by the opposite polarization (Figure 22-2). At the peak of an action potential, the membrane potential can be as much as +50 mV (inside-positive), a net change of ~120 mV. As we shall see in greater detail in Section 22.2, an action potential moves along the axon to the axon terminus at speeds of up to 100 meters per second. In humans, for instance, axons may be more than a meter long, yet it takes only a few milliseconds for an action potential to move along their length. Neurons can fire repeatedly after a brief recovery period, for example, every 4 milliseconds (ms), as in Figure 22-2. After the action potential passes through a section of a neuron, channel proteins and pumps restore the inside-negative resting potential (repolarization). The restoration process chases the action potential down the axon to the terminus, leaving the neuron ready to signal again.
EXPERIMENTAL FIGURE 22-2Recording of an axonal membrane potential over time reveals the amplitude and frequency of action potentials. An action potential is a sudden, transient depolarization of the membrane, followed by repolarization to the resting potential of about –70 mV. The axonal membrane potential can be measured with a small electrode placed into it (see Figure 11-19). This recording shows the neuron generating one action potential about every 4 milliseconds.
Page 1028
Importantly, action potentials are “all or none.” Once the threshold to start one is reached, a full firing occurs. The signal information is therefore carried primarily not by the intensity of the action potentials, but by the timing and frequency of them.
Some excitable cells are not neurons. Muscle contraction is triggered by motor neurons that synapse directly on excitable muscle cells (see Figure 22-1b). Insulin secretion from the β-islet cells of the pancreas is triggered by neurons. In both cases the activating event involves an opening of plasma membrane channels that causes changes in the transmembrane flow of ions and in the electrical properties of the regulated cells.