Multiple Molecular Mechanisms Contribute to Synaptic Plasticity
In considering how experience can change synaptic strength, it is useful to think about the structure of the chemical synapse and the process of synaptic transmission described in Section 22.3. Long-lasting changes in plasticity have been shown to involve presynaptic changes in neurotransmitter release, trans-synaptic adhesion, and postsynaptic responses to neurotransmitter. We will briefly touch on pre- and trans-synaptic mechanisms and then delve into slightly more detail with postsynaptic mechanisms, which have been studied in greater depth.
Experiences that stimulate hippocampal neurons trigger elevations in intracellular calcium, which in turn activates kinases that phosphorylate synapsins, the molecules that organize synaptic vesicles into discrete pools within the presynaptic compartment. This phosphorylation of synapsin increases the number of synaptic vesicles available for release, thereby increasing the amount of neurotransmitter that is released with a given stimulus. Experience also activates kinases that phosphorylate RIM, the molecule that tethers voltage-gated Ca2+ channels to the release machinery, and this phosphorylation is required for LTP of hippocampal synapses.
Experience-dependent changes in trans-synaptic molecules can also affect plasticity. Hippocampal learning tasks have been shown to increase the addition of polysialic acid moieties to the Neural Cell Adhesion Molecule (NCAM) at synapses. Increased polysialylation of NCAM decreases its homophilic adhesion, which is thought to be necessary for new synaptic remodeling and growth.
Synaptic plasticity also depends on activation of kinases in the postsynaptic compartment. Influx of Ca2+, through voltage-gated Ca2+ channels and specific glutamate receptors in the postsynaptic membrane, activates one particularly important kinase, the calcium-calmodulin-dependent kinase IIα, (CamKIIα, see Chapter 15 for discussion of calcium calmodulin signaling). This kinase has the special property that once activated, it can remain persistently activated even in the absence of stimulation. This is because once activated, CamKIIα autophosphorylates itself, which renders it constitutively active for about 30 minutes, during which time the kinase phosphorylates many substrates in the postsynaptic compartment, including glutamate receptors. Phosphorylation of glutamate receptors regulates their conductance and their localization, as described below. Mice lacking CamKIIα have deficits in both LTP in the hippocampus and in memory formation.
Page 1073
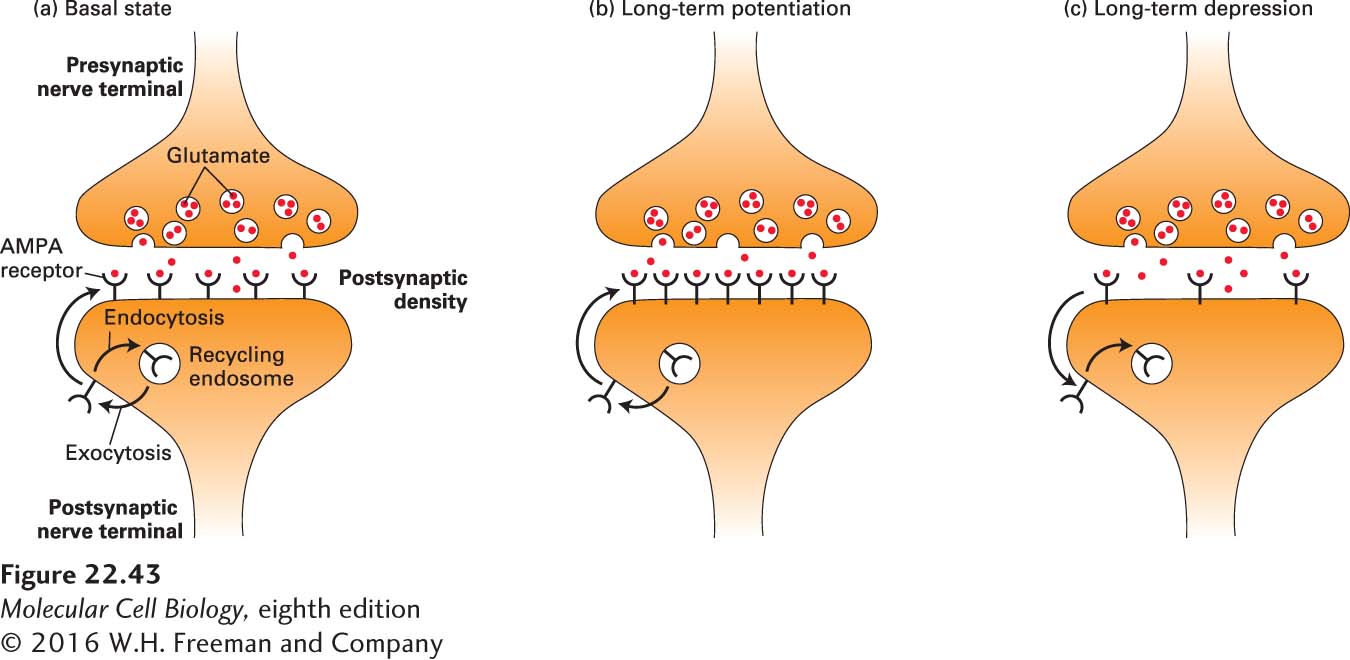
As discussed in Chapter 15, the sensitivity of a cell to external signals is determined by the number of surface receptors. In line with this concept, one of the best-characterized mechanisms underlying synaptic plasticity involves activity-dependent changes in the number of glutamate receptors that are present in the postsynaptic membrane. This process has been especially well studied in the context of hippocampal LTP and LTD (Figure 22-43). One of the major classes of glutamate receptors, called AMPA receptors, traffic constitutively to and from the plasma membrane via recycling endosomes. AMPA receptors are delivered by exocytosis at extrasynaptic sites and then laterally diffuse into the postsynaptic density, the protein-dense region of the postsynaptic site that faces the synaptic cleft and thus receives neurotransmitters released by the presynaptic terminal. AMPA receptors are removed by endocytosis, which occurs when the receptors diffuse laterally to extrasynaptic sites and are then internalized by clathrin-mediated, dynamin-dependent endocytosis.
FIGURE 22-43AMPA glutamate receptor trafficking during hippocampal LTP and LTD. (a) In the basal state, AMPA receptors (black) traffic constitutively to and from the plasma membrane of the postsynaptic compartment via recycling endosomes. Receptors are delivered to the plasma membrane lateral to the postsynaptic density via exocytosis, and are internalized by clathrin-mediated endocytosis into recycling endosomes. In the postsynaptic density, the AMPA receptors are stabilized by interactions with proteins, including transmembrane AMPA receptor regulatory proteins (TARPS, not shown). (b) Following induction of long-term potentiation (LTP) at glutamatergic synapses, there is an increase in the exocytosis of AMPA receptors and an increase in their diffusion into the postsynaptic density. This results in an increase in the number of AMPA receptors on the postsynaptic membrane, and an increase in the postsynaptic response to a given amount of glutamate release from the presynaptic neuron. (c) Following induction of long-term depression (LTD) of glutamatergic synapses, there is an increase in the diffusion of AMPA receptors out of the postsynaptic density and in their internalization into recycling endosomes. This results in a decrease in the number of AMPA receptors on the postsynaptic membrane, and a decrease in the postsynaptic response to a given amount of glutamate release from the presynaptic neuron. Regulated trafficking of AMPA receptors provides one molecular mechanism underlying the activity-dependent changes in synaptic strength that accompany synaptic plasticity and memory. See J. D. Shepherd and R. L. Huganir, 2007, Ann. Rev. Cell Dev. Biol.23:613–643.
While AMPA receptor trafficking occurs under basal conditions, it is modulated by activity through changes in actin and myosin dynamics as well as AMPA receptor interactions with scaffolding proteins and accessory subunits. One of these accessory subunits, Stargazin, mediates the interaction between AMPA receptors and the postsynaptic density protein PSD95. This interaction is critical to the localization of AMPA receptors at the synapse, since interaction with PSD95 stabilizes AMPA receptor localization within the postsynaptic density. Activity causes the phosphorylation of Stargazin, decreasing the mobility of AMPA receptors and increasing their concentrations at the synapse. Blocking Stargazin phosphorylation or dephosphorylation blocks hippocampal LTP and LTD, respectively. Stargazin is one of a family of transmembrane AMPA receptor regulatory proteins (TARPs). TARPs bind to all AMPA receptor subunits, are differentially expressed throughout the brain, and mediate the delivery of AMPA receptors to the surface and synapse of neurons.