Toll-Like Receptors Perceive a Variety of Pathogen-Derived Macromolecular Patterns
An important part of the innate immune system is its ability to immediately detect the presence of a microbial invader and respond to it. This response includes direct elimination of the invader, but it also prepares the mammalian host for a proper adaptive immune response, particularly through activation of professional APCs. These APCs are positioned throughout the epithelia (airways, gastrointestinal tract, genital tract), where contact with pathogens is most likely to occur. In the skin, a network of dendritic cells called Langerhans cells makes it virtually impossible for a pathogen that breaches this barrier to avoid contact with these professional APCs. Dendritic cells and other professional APCs detect the presence of bacteria and viruses through members of the Toll-like receptor (TLR) family. These proteins are named after the Drosophila protein Toll because of the structural and functional homology between them. Drosophila Toll was discovered because of its important role in dorsal/ventral patterning in the fruit fly, but related receptors are now recognized as capable of triggering an innate immune response in insects as well as in vertebrates.
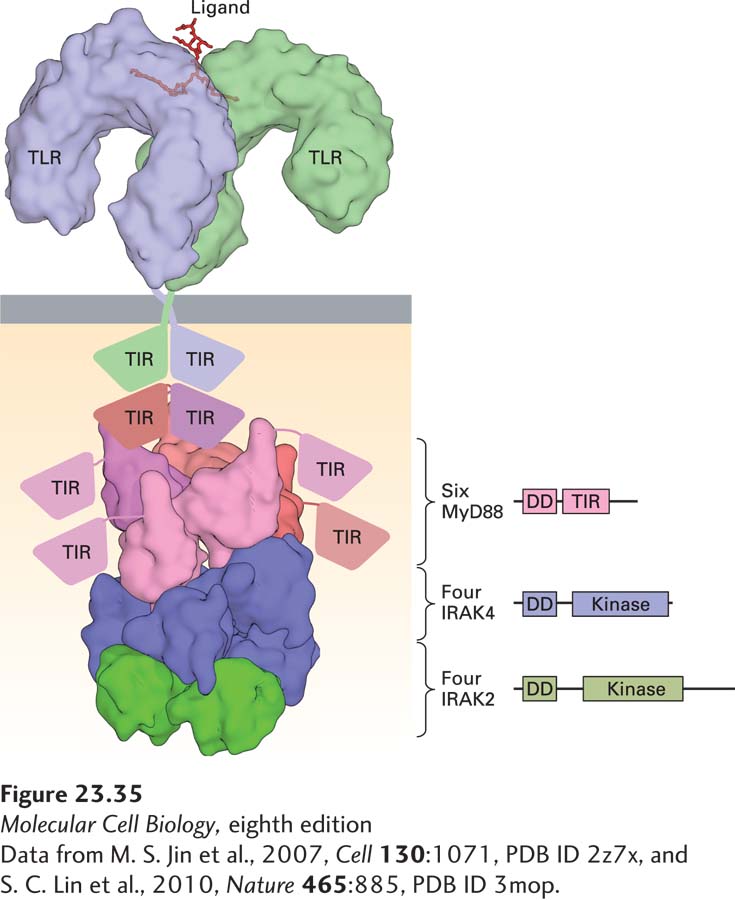
TLR Structure Toll itself and all TLRs possess a sickle-shaped extracellular domain, composed of leucine-rich repeats, that is involved in ligand recognition. The cytoplasmic portion of a TLR contains a domain responsible for the recruitment of adapter proteins to enable signal transduction. The signaling pathways engaged by TLRs have many of the same components (and outcomes) as those used by receptors for the cytokine IL-1 (Figure 23-35).
The Drosophila Toll protein interacts with its ligand, Spaetzle, itself the product of a proteolytic conversion initiated by components of the cell walls of fungi that prey on Drosophila. In the fly, activation of Toll unleashes a signaling cascade that ultimately controls the transcription of genes that encode antimicrobial peptides. The activated receptor at the cell surface communicates with the transcriptional apparatus by means of a series of adapter proteins that activate downstream kinases interposed between the TLRs and the transcription factors that are activated by them. A key step is the ubiquitin-dependent proteasomal degradation of the Cactus protein. Its removal allows the protein Dif to enter the nucleus and initiate transcription. This pathway is highly homologous in its operation and structural composition to the NF-κB pathway in mammals (see Figure 16-35).
Page 1126
[Data from M. S. Jin et al., 2007, Cell130:1071, PDB ID 2z7x, and S. C. Lin et al., 2010, Nature465:885, PDB ID 3mop.]
FIGURE 23-35Toll-like receptor activation. The extracellular portions of TLRs recognize ligands of diverse chemical nature (nucleic acids, lipopolysaccharides). The cytoplasmic portions of the TLRs, called TIR (Toll/IL1β receptor homology) domains, associate with the adapter protein MyD88, present in six copies per complex, and recruit two types of kinases, both members of the IRAK family. These complex interactions are maintained by TIR domains and death domains (DD) as shown in the figure. The assembled complex on the cytoplasmic side is referred to as the myddosome. See J. Y. Kang and J.-O. Lee, 2011, Annu. Rev. Biochem. 80:917.
[Data from M. S. Jin et al., 2007, Cell130:1071, PDB ID 2z7x, and S. C. Lin et al., 2010, Nature465:885, PDB ID 3mop.]
Diversity of TLRs There are approximately a dozen mammalian TLRs that can be activated by various microbial products and are expressed by a variety of cell types. TLR function is crucial for the activation of dendritic cells and macrophages. Neutrophils also express TLRs. The microbial products recognized by TLRs include macromolecules found in the cell envelopes of bacteria, such as lipopolysaccharides, flagellins (subunits of bacterial flagella), and bacterial lipopeptides. Direct binding of at least some of these macromolecules to TLRs has been demonstrated in crystallographic analyses of the relevant complexes. The presence of distinct classes of microbial molecules is sensed by distinct TLRs: for example, TLR4 for lipopolysaccharides; heterodimers of TLR1 and 2 and TLR2 and 6 for lipopeptides; and TLR5 for flagellin. Recognition of all bacterial envelope components occurs at the cell surface.
A second set of TLRs—TLR3, TLR7, and TLR9—sense the presence of pathogen-derived nucleic acids. They do so not at the cell surface, but rather within the endosomal compartments where these receptors reside. Mammalian DNA is methylated at many CpG dinucleotides, whereas microbial DNA generally lacks this modification. TLR9 is activated by unmethylated, CpG-containing microbial DNA. Similarly, double-stranded RNA molecules present in some virus-infected cells lead to activation of TLR3. Finally, TLR7 responds to the presence of certain single-stranded RNAs. Thus the full set of mammalian TLRs allows the recognition of a variety of macromolecules that are diagnostic for the presence of bacterial, viral, or fungal pathogens and parasites such as malaria.
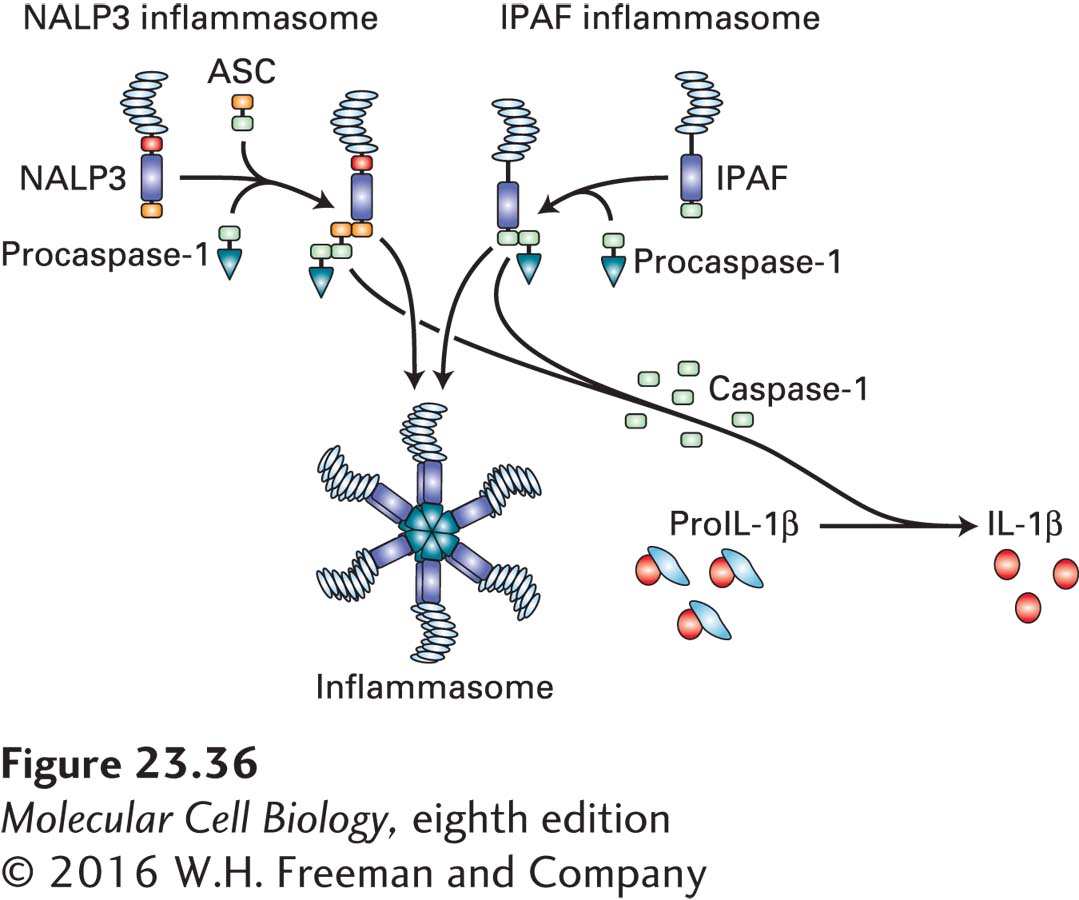
Inflammasome A variety of intracellular receptors for RNA and DNA that recognize viral RNA and are structurally distinct from TLRs have been described. The list of cytoplasmic receptors capable of recognizing DNA, both pathogen derived and host DNA derived, continues to grow. Several of these receptors participate in the assembly of the inflammasome (Figure 23-36), whose major function is the conversion of the enzyme precursor procaspase-1 to the active caspase-1. Caspase-1 is a protease that converts pro-IL-1β into active IL-1β, a cytokine that elicits a strong inflammatory response. The core components of inflammasomes are proteins with leucine-rich repeats, members of the neuronal inhibitors of apoptosis (NALP) family of proteins, and the NOD proteins, so named because of the presence of a nucleotide oligomerization domain. Ipaf-1, a protein related to the Apaf-1 molecule involved in apoptosis (see Chapter 21), allows the recruitment of an adapter protein, ASC, to mediate complex formation with procaspase-1. Assembly of this multisubunit complex allows the conversion of procaspase-1 to active caspase-1 and of pro-IL-1β to IL-1β. Many seemingly unrelated substances can induce assembly and activation of an inflammasome, including silica, uric acid crystals, and asbestos particles. Accordingly, inhibition of the inflammasome signaling cascade, or blocking of the receptor for IL-1β, has shown therapeutic promise for a variety of inflammatory diseases.
FIGURE 23-36The inflammasome. The inflammasome is a type of complex that senses the presence of cytoplasmic pathogen-derived nucleic acids and can also be activated by other danger signals, including particulate matter such as uric acid crystals or even asbestos. There are close to two dozen proteins that can participate in the formation of these complexes to yield inflammasomes of different composition, two of which are represented here schematically. Ultimately, the fully assembled inflammasome activates caspase-1, the enzyme that converts pro-IL-1β into the active, cleaved cytokine IL-1β. NALP3 = a member of the protein family characterized by the presence of NACHT, LRR, and PYD domains; ASC = apoptosis-associated Speck-like protein containing a CARD (caspase recruitment domain).
Page 1127
TLR Signaling Cascade As shown in Figure 23-35, engagement of mammalian TLRs leads to recruitment of the adapter protein MyD88, which in turn allows the binding and activation of IRAK (interleukin 1 receptor-associated kinase) proteins. After IRAK phosphorylates TNF-receptor–associated factor 6 (TRAF6), several downstream kinases come into play, leading to release of active NF-κB, a transcription factor, for translocation from the cytoplasm to the nucleus, where NF-κB activates various target genes (see Figure 16-35). These target genes include those encoding IL-1β and IL-6, which contribute to inflammation, as well as the genes for TNF and IL-12. Expression of type I interferons, small proteins with antiviral effects, is also turned on in response to TLR signaling.
Cell responses to TLR signaling are quite diverse. For professional APCs, these responses include not only production of cytokines but also the up-regulation of co-stimulatory molecules, the surface proteins important for full activation of T cells that have yet to encounter antigen (referred to as naive T cells). TLR signaling allows dendritic cells to migrate from where they encounter a pathogen to the lymph nodes draining that area, where they can interact with naive lymphocytes. Not all activated TLRs evoke an identical response. In dendritic cells, each activated TLR controls production of a particular set of cytokines. For each engaged TLR, the combination of costimulatory molecules and the cytokine profile induced by TLR engagement creates a unique activated-dendritic-cell phenotype. The identity of the microbial antigen encountered by a dendritic cell determines the pattern of the TLRs that will be activated. This pattern, in turn, shapes the differentiation pathways of activated dendritic cells, influencing the cytokines produced, the surface molecules displayed, and the chemotactic cues to which the dendritic cells respond. The mode of activation of a dendritic cell and the cytokines it produces in response create a unique local microenvironment in which T cells differentiate. Within this microenvironment, the neighboring T cells acquire the functional characteristics required to fight the infectious agent that led to engagement of the TLRs in the first place.