Inflammation Is a Complex Response to Injury That Encompasses Both Innate and Adaptive Immunity
When a vascularized tissue (one that is supplied with blood vessels) is injured, the stereotypical response that follows is inflammation. The injury may be a consequence of physical or chemical processes, such as torn muscles, a simple paper cut, or infection with a pathogen. Inflammation, also called the inflammatory response, is characterized by four classic signs: redness, swelling, heat, and pain. These signs are caused by increased leakiness of blood vessels (vasodilation), attraction of immune-system cells to the site of damage, and the production of soluble mediators of inflammation, which are responsible for the sensation of heat and pain. Inflammation provides immediate protection through the activation of the cell types and soluble products that together mount the innate immune response and create a local environment conducive to the initiation of the adaptive immune response. If it is not properly controlled, however, inflammation can also be a major cause of tissue damage.
Page 1087
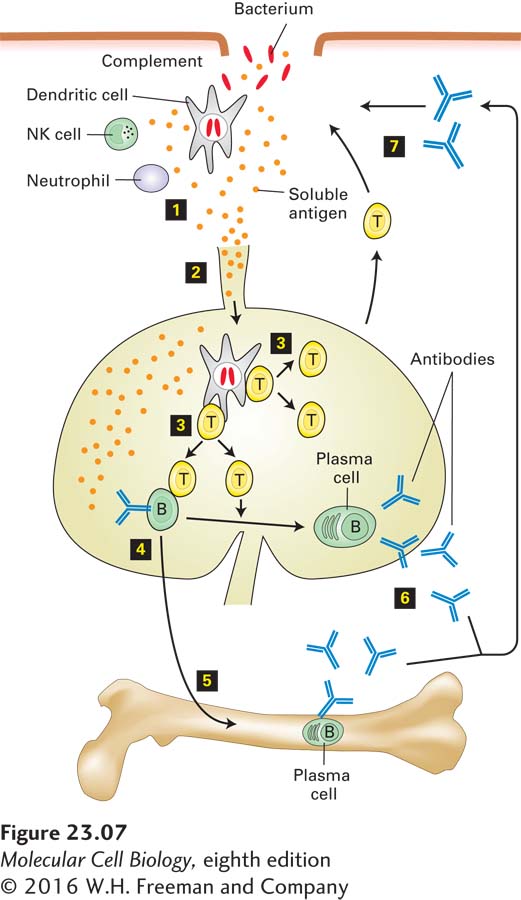
Figure 23-7 depicts the key players in the inflammatory response to bacterial pathogens and the subsequent initiation of an adaptive immune response. Tissue-resident dendritic cells sense the presence of pathogens via their TLRs and respond by releasing small soluble proteins such as cytokines and chemokines; the latter act as chemoattractants for immune-system cells. Neutrophils leave the circulation and migrate to the site of injury or infection in response to the cytokines and chemokines produced there (see Figure 20-40). Neutrophils, which constitute almost half of all circulating leukocytes, are phagocytic (see Figure 17-19), directly ingesting and destroying pathogenic bacteria and fungi. Neutrophils can interact with a wide variety of pathogen-derived macromolecules via their TLRs. Engagement of these receptors, described in detail below, activates the neutrophils, which produce more cytokines and chemokines; the latter can attract more leukocytes—neutrophils, macrophages, and ultimately lymphocytes (T and B cells)—to the area to fight the infection. Activated neutrophils can release bacteria-destroying enzymes (e.g., lysozyme and proteases) as well as small peptides with microbicidal activity, collectively called defensins. Activated neutrophils also turn on enzymes that generate the superoxide anion radical and other reactive oxygen species (see Chapter 12, pages 547-548), which can kill microbes at short range. Another cell type that contributes to the inflammatory response is tissue-resident mast cells. When activated by a variety of physical or chemical stimuli, mast cells release histamine, a small molecule that binds to G protein–coupled receptors. This binding leads to increased vascular permeability and thereby facilitates access to the tissue by plasma proteins (such as complement) that can act against the invading pathogen.
FIGURE 23-7Interplay of innate and adaptive immune responses to a bacterial pathogen. Once a bacterium breaches the host’s mechanical and chemical defenses, the bacterium is exposed to components of the complement cascade, as well as to innate immune-system cells that confer immediate protection (step 1). Various inflammatory proteins induced by tissue damage contribute to a localized inflammatory response. Local destruction of the bacterium results in the release of bacterial antigens, which are delivered, via the afferent lymphatic vessels that drain the tissue, to the lymph nodes (step 2). Dendritic cells acquire antigen at the site of infection, become migratory, and move to the lymph nodes, where they activate T cells (step 3). In the lymph nodes, antigen-stimulated T cells proliferate and acquire effector functions, including the ability to help B cells (step 4), some of which may move to the bone marrow and complete their differentiation into plasma cells there (step 5). In later stages of the immune response, activated T cells provide additional assistance to antigen-experienced B cells to yield plasma cells that secrete antigen-specific antibodies at a high rate (step 6). Antibodies produced as a consequence of the initial exposure to the bacterium act in synergy with complement to eliminate the infection (step 7), should it persist, or afford rapid protection in the case of re-exposure to the same pathogen.
A very important early response to infection or injury is the activation of a variety of plasma proteases, including the proteins of the complement cascade discussed above. As we have seen, the cleavage fragments produced during activation of these proteases attract neutrophils to the site of tissue damage (see Figure 23-5). They further induce production of cytokines such as interleukins 1 and 6 (IL-1 and IL-6), which cause inflammation. The recruitment of neutrophils also depends on an increase in vascular permeability, which is controlled in part by lipid signaling molecules (e.g., prostaglandins and leukotrienes) that are derived from phospholipids and fatty acids. All of these events occur rapidly, starting within minutes of injury. A failure to resolve the cause of this immediate response may result in chronic inflammation with ensuing tissue damage, in which cells of the adaptive immune system play an important role.
When the pathogen burden at the site of tissue damage is high, it may exceed the capacity of innate defense mechanisms to deal with the infection. Moreover, some pathogens have acquired, in the course of evolution, tools to disable or bypass innate immune defenses. In such situations, the adaptive immune response is required to help control the infection. This response depends on specialized cells that straddle the interface between adaptive and innate immunity, including macrophages and dendritic cells, which are capable of ingesting and killing pathogens as well as presenting antigens to the adaptive immune system. Dendritic cells, in particular, can initiate an adaptive immune response by delivering newly acquired pathogen-derived antigens to secondary lymphoid organs (see Figure 23-7).