Inappropriate Production of Nuclear Transcription Factors Can Induce Transformation
Mutations that create oncogenes or damage tumor-suppressor genes eventually cause changes in gene expression. These changes can be measured by comparing the amounts of different mRNAs produced in normal cells and in tumor cells. Since the most direct effect on gene expression is exerted by transcription factors, it is not surprising that many oncogenes encode transcription factors. Two examples are JUN and FOS, which initially were identified in transforming retroviruses and later found to be overexpressed in some human tumors. The JUN and FOS proto-oncogenes encode proteins that sometimes associate to form a heterodimeric transcription factor, called AP1, that binds to a sequence found in promoters and enhancers of many genes (see Figure 9-31a and Chapter 16). These proteins function as oncoproteins by activating the transcription of key genes that encode growth-promoting proteins or by inhibiting the transcription of growth-repressing genes.
Page 1161
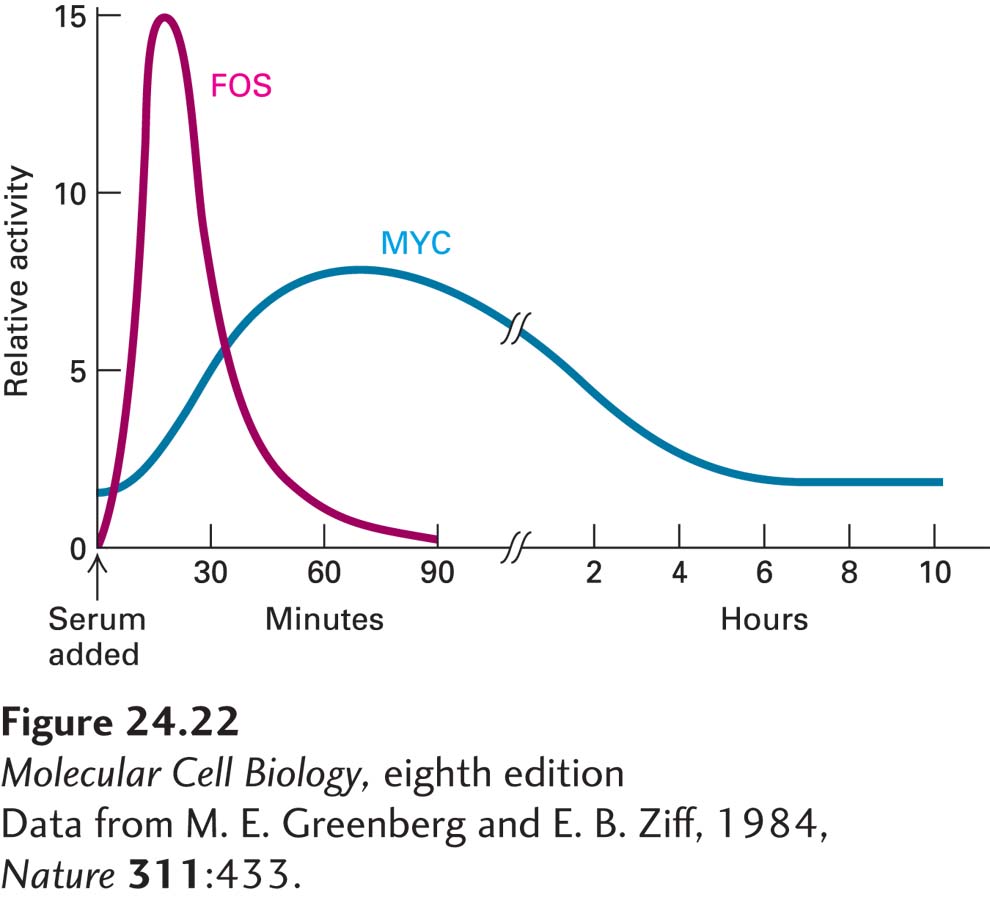
Many nuclear proto-oncogene proteins are produced when normal cells are stimulated to grow, indicating their direct role in growth control. For example, PDGF treatment of quiescent mouse 3T3 cells induces an approximately 50-fold increase in the production of the transcription factors FOS and MYC, the products of the FOS and MYC proto-oncogenes. Initially, there is a transient rise of FOS and later a more prolonged rise of MYC (Figure 24-22). The levels of both proteins decline within a few hours, a regulatory effect that may, in normal cells, help to avoid cancer. The FOS and MYC proteins stimulate transcription of genes encoding proteins that promote progression through the G1 phase of the cell cycle and the G1-to-S transition.
[Data from M. E. Greenberg and E. B. Ziff, 1984, Nature311:433.]
EXPERIMENTAL FIGURE 24-22Addition of serum to quiescent 3T3 cells yields a marked increase in the activity of two proto-oncogene products, FOS and MYC. Serum contains factors such as platelet-derived growth factor (PDGF) that stimulate the growth of quiescent cells. One of the earliest effects of growth factors is to induce expression of FOS and MYC, whose encoded proteins are transcription factors.
[Data from M. E. Greenberg and E. B. Ziff, 1984, Nature311:433.]
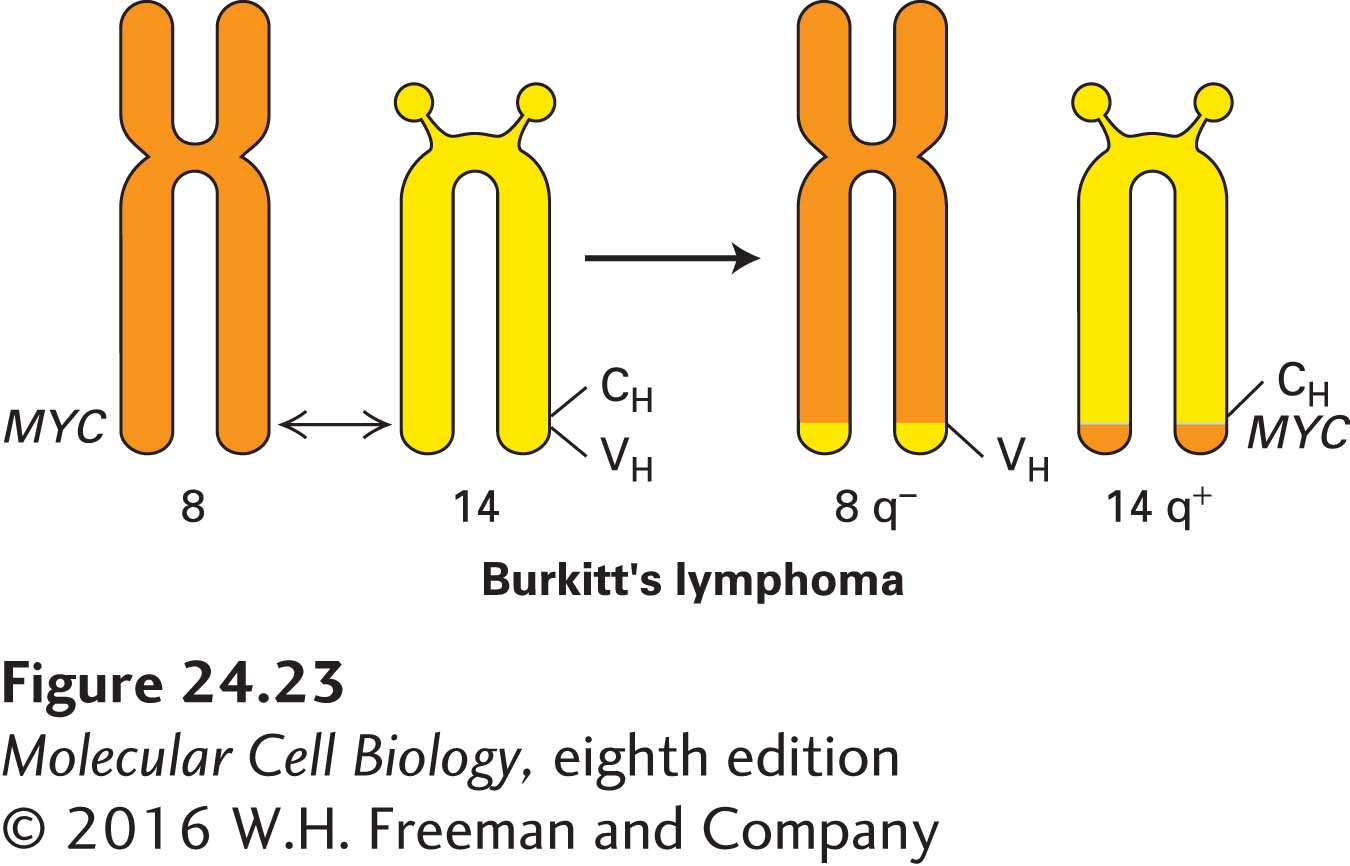
In normal cells, FOS and MYC mRNAs and the proteins they encode are intrinsically unstable and degrade rapidly after the genes are transcribed. Some of the genetic changes that turn FOS from a normal gene into an oncogene involve deletions of the sequences that normally make the FOS mRNA and protein short-lived. Conversion of the MYC proto-oncogene into an oncogene can occur by different mechanisms. In cells of the human tumor known as Burkitt’s lymphoma, the MYC gene is translocated to a site near the heavy-chain antibody genes, which are normally active in antibody-producing white blood cells (Figure 24-23). The MYC translocation is a rare aberration of the normal DNA rearrangements that occur during maturation of antibody-producing cells. The translocated MYC gene, now regulated by the antibody-gene enhancer, is continually highly expressed, causing the cell to become cancerous. Localized amplification of a segment of DNA containing the MYC gene, which occurs in several human tumors, also causes inappropriately high production of the otherwise normal MYC protein.
FIGURE 24-23Chromosomal translocation in Burkitt’s lymphoma. As a result of a translocation between chromosomes 8 and 14, the MYC gene is placed adjacent to the gene for part of the antibody heavy chain (CH), leading to overproduction of the MYC transcription factor in lymphocytes and hence their growth into a lymphoma.
The MYC gene encodes a basic helix-loop-helix protein that acts as part of a set of interacting proteins that can dimerize in various combinations, bind to DNA, and coordinately regulate the transcription of target genes. Other members of this protein set include MAD, MAX, and MNT. MAX can heterodimerize with MYC, MAD, and MNT. MYC-MAX dimers regulate genes that control proliferation, such as cyclins. MAD proteins inhibit MYC proteins, which has led to an interest in using MAD proteins, or drugs that stimulate MAD proteins, to rein in excessive MYC activity that contributes to tumor formation. MYC protein complexes affect transcription by recruiting chromatin-modifying complexes containing histone acetyl transferases (which usually stimulate transcription; see Chapter 9) to MYC target genes. MAD and MNT work with the SIN3 co-repressor protein to bring in histone deacetylases that help to block transcription. Together, all these proteins form a regulatory network that employs protein-protein association, variations in DNA binding, and transcriptional regulation to control cell proliferation. Overproduction of MYC protein tips the scales in favor of cell growth and division.