Cryoelectron Microscopy Allows Visualization of Specimens Without Fixation or Staining
Standard transmission electron microscopy cannot be used to study live cells, and the absence of water in samples causes macromolecules to become denatured and nonfunctional. However, hydrated, unfixed, and unstained biological specimens can be viewed directly in a transmission electron microscope if the samples are frozen. In cryoelectron microscopy, an aqueous suspension of a sample is applied to a grid in an extremely thin film, frozen in liquid nitrogen, and maintained in this state by means of a special mount. The frozen sample is then placed in the electron microscope. The very low temperature (−196 °C) keeps water from evaporating, even in a vacuum. Thus the sample can be observed in detail in its native, hydrated state without fixing or heavy metal staining. By computer-based averaging of hundreds of images, a three-dimensional model can be generated almost to atomic resolution. For example, this method has been used to generate models of ribosomes, the muscle calcium pump discussed in Chapter 11, and the three-dimensional structure of a poliovirus capsid (see Figure 5-43b).
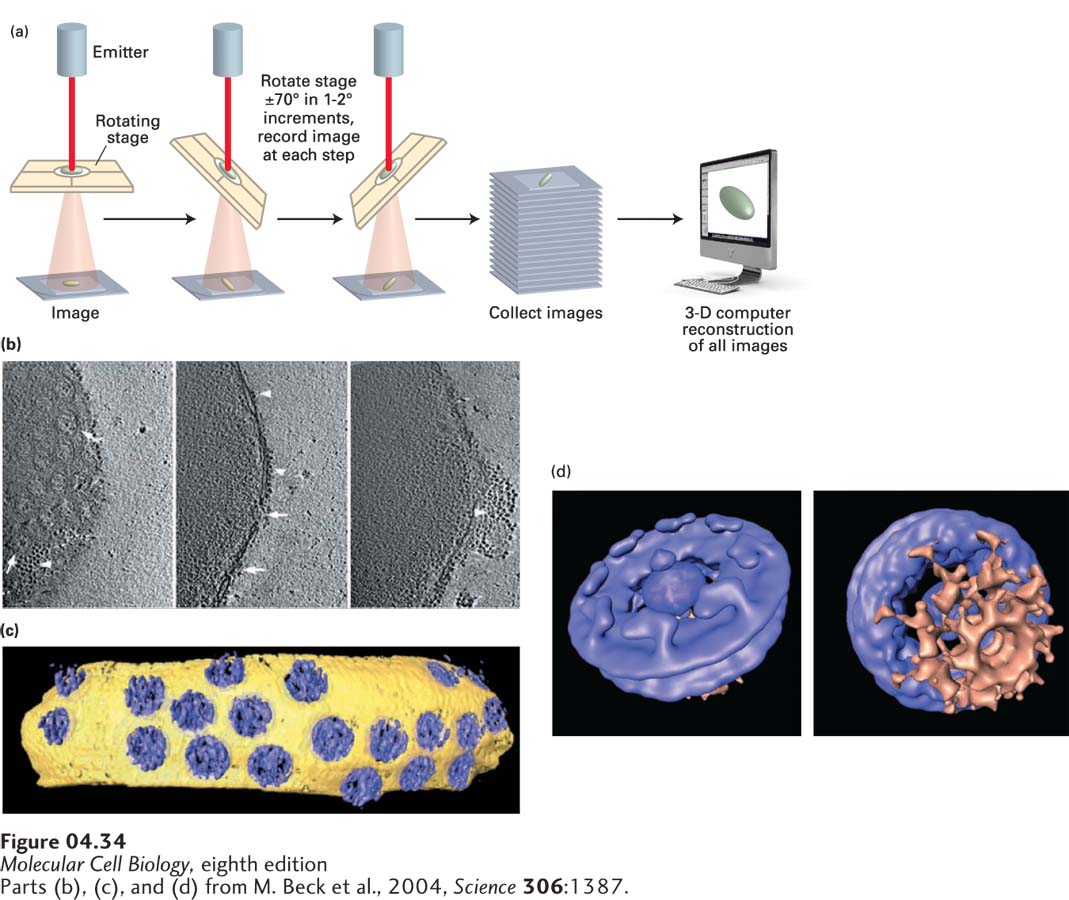
An extension of this technique, cryoelectron tomography, allows researchers to determine the three-dimensional architecture of organelles or even whole cells embedded in ice; that is, in a state close to life. A single picture is a two-dimensional representation of a structure that lacks depth information. However, looking at the same structure from different angles gives us a three-dimensional perspective. In cryoelectron tomography, the specimen holder is tilted in small increments around the axis perpendicular to the electron beam; thus images of the object viewed from different directions are obtained (Figure 4-34a, b). The images are then merged computationally into a three-dimensional reconstruction termed a tomogram (Figure 4-34c, d). A disadvantage of cryoelectron tomography is that the samples must be relatively thin, about 200 nm; this is much thinner than the samples (200 µm thick) that can be studied by confocal light microscopy.
Page 161
[Parts (b), (c), and (d) from M. Beck et al., 2004, Science306:1387.]
FIGURE 4-34Structure of the nuclear pore complex (NPC) imaged by cryoelectron tomography. (a) In cryoelectron tomography, a semicircular series of two-dimensional projection images is recorded from the three-dimensional specimen that is located at the center; the specimen is tilted while the electron optics and detector remain stationary. The three-dimensional structure is then computed from the collected two-dimensional images. (b) Isolated nuclei from the cellular slime mold Dictyostelium discoideum were quick-frozen in liquid nitrogen and maintained in this state as the sample was observed in the electron microscope. The panel shows three sequential tilted images. Different orientations of NPCs (arrows) are shown in top view (left and center) and side view (right). Ribosomes connected to the outer nuclear membrane are visible, as is a patch of rough ER (arrowheads). (c) Computer-generated surface-rendered representation of a segment of the nuclear envelope membrane (yellow) studded with NPCs (blue). (d) By averaging the images of multiple nuclear pores, much more detail can be discerned. See S. Nickell et al., 2006, Nature Rev. Mol. Cell Biol.7:225.
[Parts (b), (c), and (d) from M. Beck et al., 2004, Science306:1387.]