Native DNA Is a Double Helix of Complementary Antiparallel Strands
The modern era of molecular biology began in 1953 when James D. Watson and Francis H. C. Crick proposed that DNA has a double-helical structure. Their proposal was based on analysis of x-ray diffraction patterns of DNA fibers generated by Rosalind Franklin and Maurice Wilkins, which showed that the structure was helical, and analyses of the base composition of DNA from multiple organisms by Erwin Chargaff and colleagues. Chargaff’s studies revealed that while the base composition of DNA (percentages of A, T, G, and C) varies greatly between distantly related organisms, the percentage of A always equals the percentage of T, and the percentage of G always equals the percentage of C, in all organisms. Based on these discoveries and the structures of the four nucleotides, Watson and Crick performed careful molecular model building, proposing a double helix, with A always hydrogen-bonded to T and G always hydrogen-bonded to C at the axis of the double helix, as the structure of DNA. The Watson and Crick model proved correct and paved the way for our modern understanding of how DNA functions as the genetic material. Today our most accurate models for DNA structure come from high-resolution x-ray diffraction studies of crystals of DNA, made possible by the chemical synthesis of large amounts of short DNA molecules of uniform length and sequence that are amenable to crystallization (Figure 5-3a).
[Part (a) data from R. Wing et al., 1980, Nature287:755, PDB ID 1bna.]
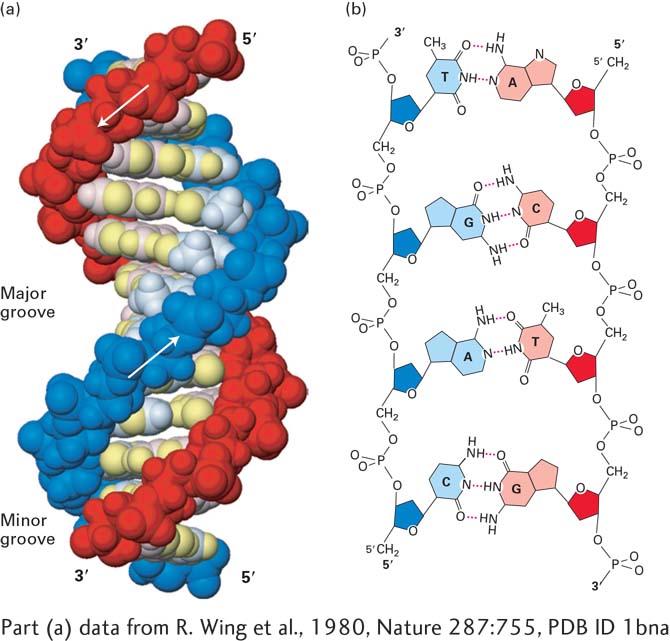
FIGURE 5-3The DNA double helix. (a) Space-filling model of B DNA, the most common form of DNA in cells. The bases (light shades) project inward from the sugar-phosphate backbones (dark red and blue) of each strand, but their edges are accessible through major and minor grooves. Arrows indicate the 5′→3′ direction of each strand. Hydrogen bonds between the bases are in the center of the structure. The major and minor grooves are lined by potential hydrogen bond donors and acceptors (highlighted in yellow). (b) Chemical structure of DNA double helix. This extended schematic shows the two sugar-phosphate backbones and hydrogen bonding between the Watson-Crick base pairs, A·T and G·C. See R. E. Dickerson, 1983, Sci. Am.249:94.
[Part (a) data from R. Wing et al., 1980, Nature287:755, PDB ID 1bna.]
Page 171
DNA consists of two associated polynucleotide strands that wind together to form a double helix. The two sugar-phosphate backbones are on the outside of the double helix, and the bases project into the interior. The adjoining bases in each strand stack on top of one another in parallel planes (see Figure 5-3a). The orientation of the two strands is antiparallel; that is, their 5′→3′ directions are opposite. The strands are held in precise register by formation of base pairs between the two strands: A is paired with T through two hydrogen bonds; G is paired with C through three hydrogen bonds (Figure 5-3b). This base-pair complementarity is a consequence of the size, shape, and chemical composition of the bases. The presence of thousands of such hydrogen bonds in a DNA molecule contributes greatly to the stability of the double helix. Hydrophobic and van der Waals interactions between the stacked adjacent base pairs further stabilize the double-helical structure.
In natural DNA, A always hydrogen-bonds with T and G with C, forming A·T and G·C base pairs as shown in Figure 5-3b. These associations, always between a larger purine and a smaller pyrimidine, are often called Watson-Crick base pairs. Two polynucleotide strands, or regions thereof, in which all the nucleotides form such base pairs are said to be complementary. However, in theory and in synthetic DNAs, other base pairs can form. For example, guanine (a purine) could theoretically form hydrogen bonds with thymine (a pyrimidine), causing only a minor distortion in the helix. The space available in the helix would also allow pairing between the two pyrimidines cytosine and thymine. Although the nonstandard G·T and C·T base pairs are not normally found in DNA, G·U base pairs are quite common in double-helical regions that form within otherwise single-stranded RNA. Nonstandard base pairs do not occur naturally in double-stranded (duplex) DNA because the DNA copying enzyme, described later in this chapter, does not permit them.
Most DNA in cells takes the form of a right-handed helix. The x-ray diffraction pattern of DNA indicates that the stacked bases are regularly spaced 0.34 nm apart along the helix axis. The helix makes a complete turn every 3.4 to 3.6 nm, depending on the sequence; thus there are about 10–10.5 base pairs per turn. This helical form, referred to as the B form of DNA, is the normal form present in most DNA stretches in cells. On the outside of the helix, the spaces between the intertwined strands form two helical grooves of different widths, described as the major groove and the minor groove (see Figure 5-3a). As a consequence, the atoms on the edges of each base within these grooves are accessible from outside the helix, forming two types of binding surfaces. DNA-binding proteins can “read” the sequence of bases in duplex DNA by contacting atoms in either the major or the minor grooves.
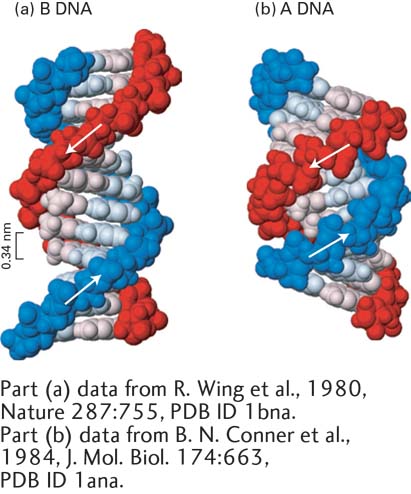
Under laboratory conditions in which most of the water is removed from DNA, the crystallographic structure of DNA changes to the A form, which is wider and shorter than B-form DNA, with a wider and deeper major groove and a narrower and shallower minor groove (Figure 5-4). RNA-DNA and RNA-RNA helices also exist in this form in cells and in vitro.
[Part (a) data from R. Wing et al., 1980, Nature287:755, PDB ID 1bna. Part (b) data from B. N. Conner et al., 1984, J. Mol. Biol.174:663, PDB ID 1ana.]
FIGURE 5-4Comparison of A-Form and B-Form DNA. The sugar-phosphate backbones of the two polynucleotide strands, which are on the outside in both structures, are shown in red and blue; the bases (lighter shades) are oriented inward. (a) The B form of DNA has about 10.5 base pairs per helical turn. Adjacent stacked base pairs are 0.34 nm apart. (b) The more compact A form of DNA has 11 base pairs per turn, with a much deeper major groove and a much shallower minor groove than B-form DNA.
[Part (a) data from R. Wing et al., 1980, Nature287:755, PDB ID 1bna. Part (b) data from B. N. Conner et al., 1984, J. Mol. Biol.174:663, PDB ID 1ana.]
Page 172
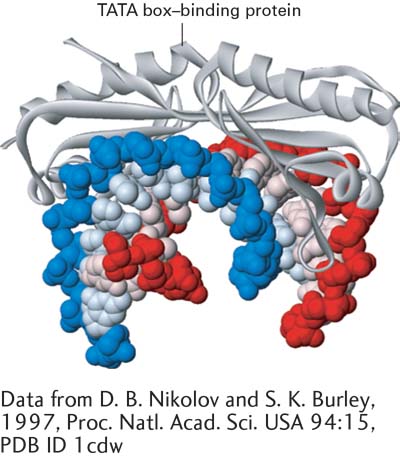
Important modifications in the structure of standard B-form DNA come about as a result of protein binding to specific DNA sequences. Although the multitude of hydrogen and hydrophobic bonds between the bases provides stability to DNA, the double helix is flexible about its long axis. Unlike the α helix in proteins (see Figure 3-4), it has no hydrogen bonds parallel to the axis of the helix. This property allows DNA to bend when complexed with a DNA-binding protein, such as the transcription factor TBP (Figure 5-5). Bending of DNA is also critical to the dense packing of DNA in chromatin, the protein-DNA complex in which nuclear DNA occurs in eukaryotic cells (see Chapter 8).
[Data from D. B. Nikolov and S. K. Burley, 1997, Proc. Natl. Acad. Sci. USA94:15, PDB ID 1cdw.]
FIGURE 5-5Interaction with a protein can bend DNA. The conserved C-terminal domain of the TATA box–binding protein (TBP) binds to the minor groove of specific DNA sequences rich in A and T, untwisting and sharply bending the double helix. Transcription of most eukaryotic genes requires participation of TBP.
[Data from D. B. Nikolov and S. K. Burley, 1997, Proc. Natl. Acad. Sci. USA94:15, PDB ID 1cdw.]
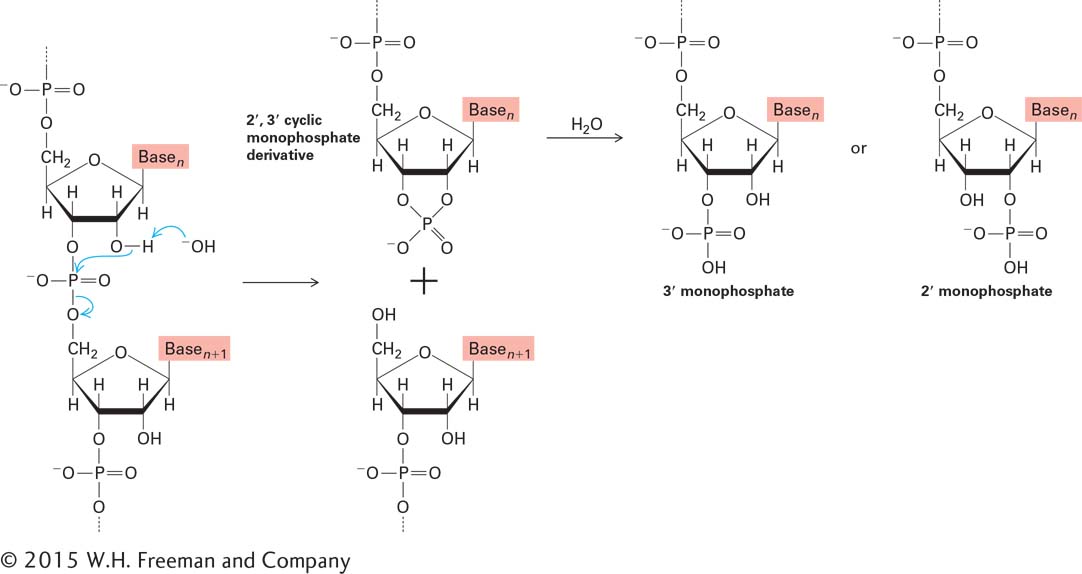
Why did DNA, rather than RNA, evolve to be the carrier of genetic information in cells? The hydrogen at the 2′ position in the deoxyribose of DNA makes it a far more stable molecule than RNA, which instead has a hydroxyl group at the 2′ position of ribose (see Figure 2-16). The 2′-hydroxyl groups in RNA participate in the slow, OH−-catalyzed hydrolysis of phosphodiester bonds at neutral pH (Figure 5-6). The absence of 2′-hydroxyl groups in DNA prevents this process. Therefore, the presence of deoxyribose in DNA makes it a more stable molecule—a characteristic that is critical to its function in the long-term storage of genetic information.
FIGURE 5-6 Base-catalyzed hydrolysis of RNA. The 2′-hydroxyl group in RNA can act as a nucleophile, attacking the phosphodiester bond. The 2′,3′ cyclic monophosphate derivative is further hydrolyzed to a mixture of 2′ and 3′ monophosphates. This mechanism of phosphodiester bond hydrolysis cannot occur in DNA, which lacks 2′-hydroxyl groups.