Homologous Recombination Can Repair DNA Damage and Generate Genetic Diversity
At one time homologous recombination was thought to be a minor repair process in human cells. This view changed when it was realized that several human cancers are potentiated by inherited mutations in genes that are essential for homologous recombination repair (see Table 24-1). For example, some women with an inherited susceptibility to breast cancer have a mutation in one allele of either the BRCA-1 or the BRCA-2 gene, both of which encode proteins participating in this repair process. Loss or inactivation of the second allele inhibits the homologous recombination repair pathway and thus tends to induce cancer in mammary or ovarian epithelial cells. Yeasts can use homologous recombination to repair double-strand breaks induced by γ-irradiation. Isolation and analysis of radiation-sensitive (RAD) mutants that are deficient in this repair system facilitated study of the process. Virtually all the yeast Rad proteins have homologs in the human genome, and the human and yeast proteins function in an essentially identical fashion.
A variety of DNA lesions that are not repaired by the mechanisms discussed earlier can be repaired by mechanisms in which the damaged DNA sequence is copied from an undamaged copy of the same or a highly homologous sequence on the homologous chromosome in diploid organisms or the sister chromosome following DNA replication in haploid and diploid organisms. These mechanisms involve an exchange of strands between separate DNA molecules and hence are referred to as recombination.
In addition to providing a mechanism for DNA repair, similar recombination mechanisms generate genetic diversity among the individuals of a species by causing the exchange of large regions of chromosomes between the maternal and paternal pair of homologous chromosomes during meiosis, the special type of cell division that generates germ cells (sperm and eggs) (see Figure 6-3). In fact, the exchange of regions of homologous chromosomes, called crossing over, is required for proper segregation of chromosomes during the first meiotic cell division. Meiosis and the consequences of generating new combinations of maternal and paternal genes by recombination are discussed further in Chapter 6, and the mechanisms leading to proper segregation of chromosomes during meiosis are discussed in Chapter 19. Here we focus on the molecular mechanisms of DNA recombination, highlighting the exchange of DNA strands between two recombining DNA molecules.
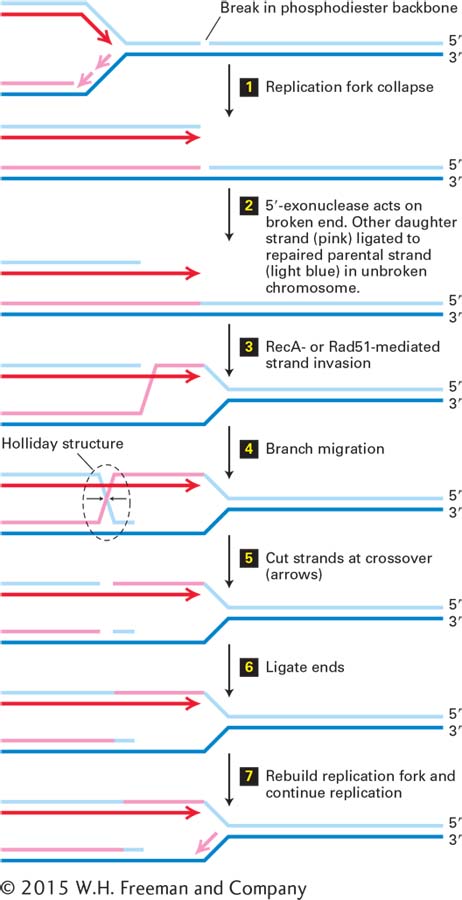
FIGURE 5-40Recombinational repair of a collapsed replication fork. Parent strands are light and dark blue. The leading daughter strand is dark red, and the lagging daughter strand pink. Diagonal lines in step 3 and beyond represent a single phosphodiester bond from the DNA strand of the corresponding color. Small black arrows following step 4 represent cleavage of the phosphodiester bonds at the crossover of DNA strands in the Holliday structure. See http://www.sheffield.ac.uk/mbb/ruva for an animation of branch migration catalyzed by E. coli proteins RuvA and RuvB. See the text for a discussion.
Repair of a Collapsed Replication Fork An example of recombinational DNA repair is the repair of a “collapsed” replication fork. If a break in the phosphodiester backbone of one DNA strand (called a “nick”) is not repaired before a replication fork passes, the replicated portions of the daughter chromosomes become separated when the replication helicase reaches the nick in the parent strand because there are no covalent bonds between the two fragments of the parent strand on either side of the nick. This process is called replication fork collapse (Figure 5-40, step 1). If the break in the double-stranded daughter DNA molecule is not repaired, it is generally lethal to at least one daughter cell following cell division because of the loss of genetic information between the break and the end of the chromosome. The recombination process that repairs the resulting double-strand break and regenerates a replication fork involves multiple enzymes and other proteins, only some of which are mentioned here.
The first step in the repair of the double-strand break is exonucleolytic digestion of the strand with its 5′ end at the broken end of DNA, leaving the strand with a 3′ end at the break single-stranded (Figure 5-40, step 2). The lagging nascent strand (pink) base-paired to the unbroken parent strand (dark blue) is ligated to the unreplicated portion of the parent chromosome (light blue), as shown in Figure 5-40, step 2. A protein required for the next step is RecA in bacteria, or the homologous Rad51 in S. cerevisiae and other eukaryotes. Multiple RecA/Rad51 molecules bind to the single-stranded DNA and catalyze its hybridization to a perfectly or nearly perfectly complementary sequence in another, homologous, double-stranded DNA molecule. The complementary strand of this target double-stranded DNA (dark blue) is displaced as a single-stranded loop of DNA over the region of hybridization to the invading strand (Figure 5-40, step 3). This RecA/Rad51–catalyzed invasion of a duplex DNA by a single-stranded complement of one of the strands is key to the recombination process. This process is called strand invasion, and because there is no change in the number of base pairs, it does not require an input of energy.
Next the hybrid region between target DNA and the invading strand is extended in the direction away from the break by proteins that use energy from ATP hydrolysis. This process is called branch migration (Figure 5-40, step 4) because the position where the target DNA strand crosses from one complementary strand (dark blue) to its complement in the broken DNA molecule (dark red)—that is, the pink diagonal line after step 3—is called a branch in the DNA structure. In this diagram, the diagonal lines represent only one phosphodiester bond. Molecular modeling and other studies show that the first base on either side of the branch is base-paired to a complementary nucleotide. As this branch migrates to the left, the number of base pairs remains constant; one new base pair formed with the (red) invading strand is matched by the loss of one base pair with the parent (blue) strand.
Page 210
When the hybrid region extends beyond the 5′ end of the broken strand that was digested by the 5′-exonuclease in step 2 (light blue), the single-stranded parent DNA strand generated (light blue) base-pairs with the complementary region of the other parent strand (dark blue), which becomes single-stranded as the branch migrates to the left (Figure 5-40, step 4). The resulting structure is called a Holliday structure, after Robin Holliday, the geneticist who first proposed it as an intermediate in genetic recombination. Again, the diagonal lines in the diagram following step 4 represent single phosphodiester bonds, and all bases in the Holliday structure are base-paired to complementary bases in the parent strands. Cleavage of the phosphodiester bonds that cross over from one parent strand to the other (step 5) and ligation of the 5′ and 3′ ends base-paired to the same parent strands (step 6) result in the generation of a structure similar to a replication fork. Rebinding of replication fork proteins results in extension of the leading strand past the point of the original strand break and re-initiation of lagging-strand synthesis (step 7), thus regenerating a replication fork. The overall process allows the ligated upper strand in the lower molecule following step 2 to serve as a template for extension of the leading strand in step 7.
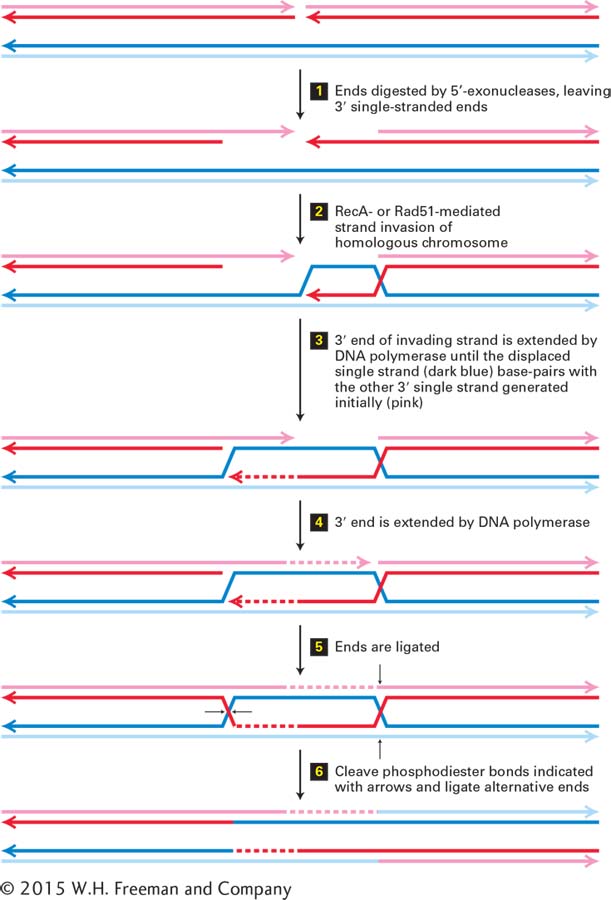
Repair of a Double-Strand Break by Homologous Recombination A similar mechanism, called homologous recombination, can repair a double-strand break in a chromosome and can also exchange large segments of two double-stranded DNA molecules (Figure 5-41). First, the broken ends of the DNA molecule are digested by 5′-exonucleases, leaving a single-stranded region of DNA with a 3′ end (step 1). RecA in bacteria or Rad51 in eukaryotes then catalyzes invasion of one of these 3′ ends into the homologous region of the homologous chromosome, as described above for repair of a collapsed replication fork (step 2). The 3′ end of the invading DNA strand is then extended by a DNA polymerase, displacing the parent strand as an enlarging single-stranded loop of DNA (dark blue) (step 3). When the loop extends to a sequence that is complementary to the other broken and 5′-exonuclease –digested end of DNA (the fragment on the left following step 1), the complementary sequences base-pair (diagram following step 3). This 3′ end is then extended by a DNA polymerase using the displaced single-stranded loop of parent DNA (dark blue) as a template (step 4).
FIGURE 5-41Repair of a double-strand break by homologous recombination. For simplicity, each DNA double helix is represented by two parallel lines with the polarities of the strands indicated by arrowheads at their 3′ ends. The upper molecule has a double-strand break. Note that in the diagram of the upper DNA molecule, the strand with its 3′ end at the right is on the top, while in the diagram of the lower DNA molecule, this strand is drawn on the bottom. See the text for discussion. See T. L. Orr-Weaver and J. W. Szostak, 1985, Microbiol. Rev.49:33.
Next the new 3′ ends are ligated (step 5) to the exonuclease-digested 5′ ends. This generates two Holliday structures in the paired molecules (step 5). Branch migration of these Holliday structures can occur in either direction (not diagrammed). Finally, cleavage of the strands at the positions shown by the arrows, and ligation of the alternative 5′ and 3′ ends at each cleaved Holliday structure, generates two recombinant chromosomes that contain the DNA of one parent DNA molecule on one side of the initial break point (pink and red strands), and the DNA of the other parent DNA molecule on the other side of the break point (light and dark blue) (step 6). The region in the immediate vicinity of the initial break point forms a heteroduplex, in which one strand from one parent is base-paired to the complementary strand of the other parent (pink or red strand base-paired to light or dark blue strand). Base-pair mismatches between the two parent strands are usually repaired by the repair mechanisms discussed above to generate a complementary base pair. In the process, sequence differences between the two parents are lost, a process referred to as gene conversion.
Page 211
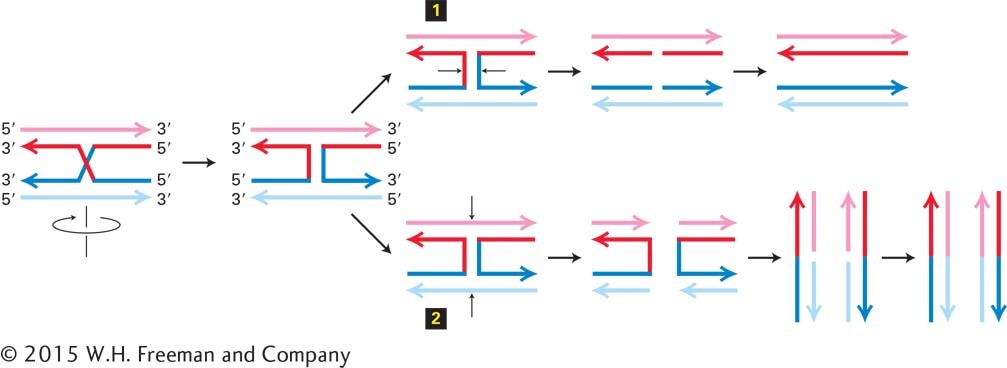
Figure 5-42 diagrams how cleavage of one or the other pair of strands at the four-way strand junction in the Holliday structure generates parent or recombinant molecules. This process, called resolution of the Holliday structure, separates DNA molecules initially joined by RecA/Rad51–catalyzed strand invasion. Each Holliday structure in the intermediate following Figure 5-41, step 5, can be cleaved and religated in the two possible ways shown by the two sets of small black arrows in Figure 5-42 (step 1 or step 2). Consequently, there are four possible products of the recombination process shown in Figure 5-41. After ligation of the cleaved ends, two of these products regenerate the parent chromosomes [with the exception of the heteroduplex region at the break point that is repaired into the sequence of one parent or the other (gene conversion)]. The other two possible products generate recombinant chromosomes as shown in Figure 5-41.
Page 212
FIGURE 5-42Alternative resolution of a Holliday structure. Diagonal and vertical lines represent a single phosphodiester bond. It is simplest to diagram the process by rotating the diagram of the bottom molecule 180° so that the top and bottom molecules have the same strand orientations. Cutting the bonds as shown in 1 and ligating the ends as indicated regenerates the original chromosomes. Cutting the strands as shown in 2 and religating as shown at the bottom generates recombinant chromosomes