Lytic Viral Growth Cycles Lead to Death of Host Cells
Although details vary among different types of viruses, those that exhibit a lytic cycle of growth proceed through the following general stages:
Adsorption—Virion interacts with a host cell by binding of multiple copies of capsid protein to specific receptors on the cell surface.
Penetration—Viral genome crosses the host plasma membrane. For some viruses, viral proteins packaged inside the capsid also enter the host cell.
Replication—Viral mRNAs are produced with the aid of the host-cell transcription machinery (DNA viruses) or by viral enzymes (RNA viruses). For both types of viruses, viral mRNAs are translated by the host-cell translation machinery. Production of multiple copies of the viral genome is carried out either by viral proteins alone or with the help of host-cell proteins.
Assembly—Viral proteins and replicated genomes associate to form progeny virions.
Release—The host cell either ruptures suddenly (lysis), releasing all the newly formed virions at once, or disintegrates gradually, releasing the virions slowly. Both types of release lead to the death of the infected cell.
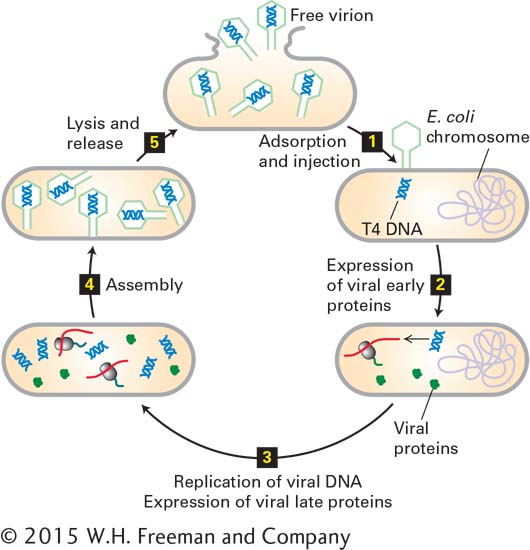
Figure 5-45 illustrates the lytic cycle for T4 bacteriophage, a nonenveloped DNA virus that infects E. coli. Viral capsid proteins generally are made in large amounts because many copies of them are required for the assembly of each progeny virion. In each infected E. coli cell, about 100–200 T4 progeny virions are produced and released by lysis.
FIGURE 5-45Lytic replication cycle of a nonenveloped bacterial virus.E. coli bacteriophage T4 has a double-stranded DNA genome and lacks a membrane envelope. After viral capsid proteins at the tip of the tail in T4 interact with specific receptor proteins on the exterior of the host cell, the viral genome is injected into the host (step 1). Host-cell enzymes then transcribe viral “early” genes into mRNAs and subsequently translate these mRNAs into viral “early” proteins (step 2). The early proteins replicate the viral DNA and induce expression of viral “late” proteins by host-cell enzymes (step 3). The viral late proteins include capsid and assembly proteins and enzymes that degrade the host-cell DNA, supplying nucleotides for synthesis of more viral DNA. Progeny virions are assembled in the cell (step 4) and released (step 5) when viral proteins lyse the cell. Newly liberated viruses initiate another cycle of infection in other host cells.
Page 214
The lytic cycle is somewhat more complicated for DNA viruses that infect eukaryotic cells. In most such viruses, the DNA genome is transported (with some associated proteins) into the cell nucleus. Once inside the nucleus, the viral DNA is transcribed into RNA by the host’s transcription machinery. Processing of the viral RNA primary transcript by host-cell enzymes yields viral mRNA, which is transported to the cytoplasm and translated into viral proteins by host-cell ribosomes, tRNA, and translation factors. The viral proteins are then transported back into the nucleus, where some of them either replicate the viral DNA directly or direct cellular proteins to replicate the viral DNA, as in the case of SV40, discussed earlier. Association of the capsid proteins with the newly replicated viral DNA occurs in the nucleus, yielding thousands to hundreds of thousands of progeny virions. Since mammalian cells are about a thousand times larger than bacterial cells, on the order of a thousand times as many virions are produced per cell. However, since many more proteins must be synthesized and viral genomes replicated to assemble this large number of virions, one cycle of infection takes much longer for an animal virus than for bacteriophage of comparable complexity. Bacteriophage T4 replicates in and lyses E. coli cells in about 20 minutes, whereas replication of poliovirus and lysis of host cells requires about 8 hours. Many animal viruses are considerably slower than poliovirus, requiring 2 days or more for lysis of an infected cell and release of progeny virions.
Page 215
Most plant and animal viruses with an RNA genome do not require nuclear functions for lytic replication. In some of these viruses, a virus-encoded enzyme that enters the host cell during penetration transcribes the viral genomic RNA into mRNA in the cell cytoplasm. The mRNA is directly translated into viral proteins by the host-cell translation machinery. One or more of these proteins then produces additional copies of the viral RNA genome. Finally, progeny genomes are associated with newly synthesized capsid proteins to form progeny virions in the cytoplasm.
After the synthesis of hundreds to hundreds of thousands of new virions has been completed, depending on the type of virus and host cell, most infected bacterial cells and some infected plant and animal cells are lysed, releasing all the virions at once. In many plant and animal viral infections, however, no discrete lytic event occurs; rather, the dead host cell releases the virions as it gradually disintegrates.
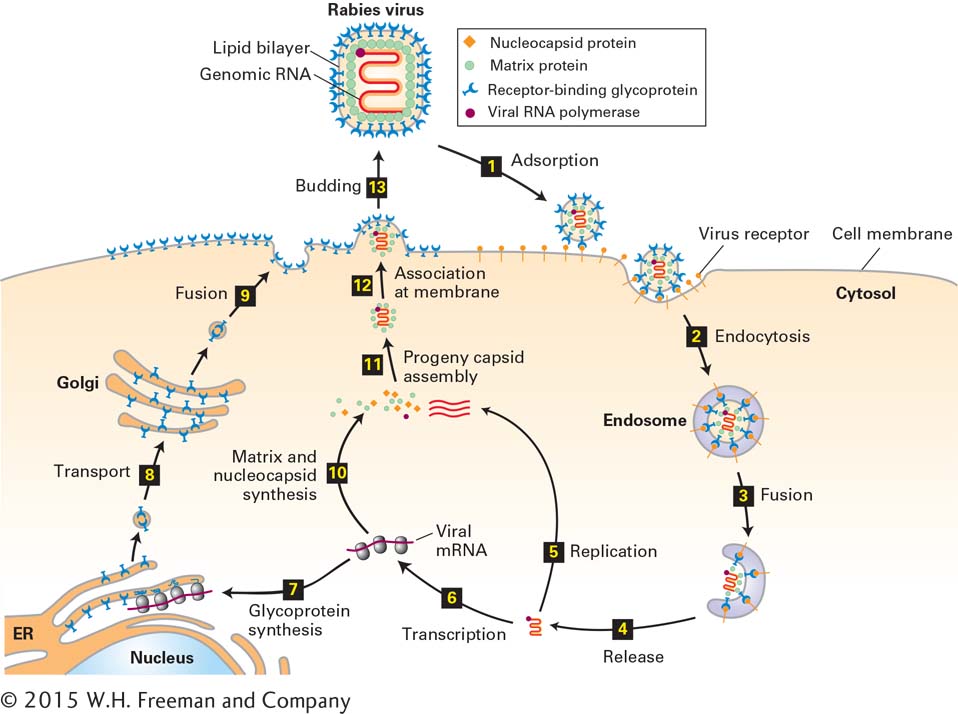
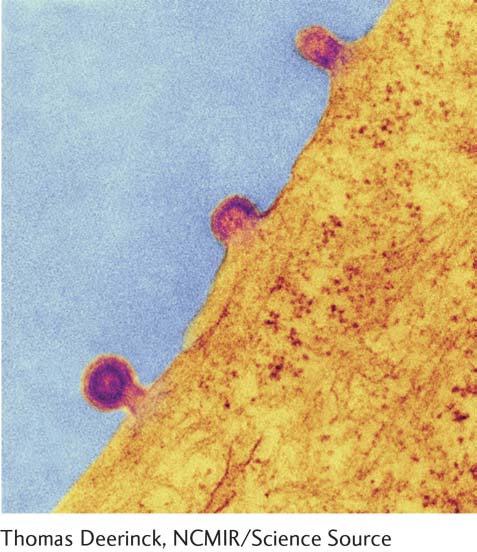
As noted previously, enveloped animal viruses are surrounded by an outer phospholipid bilayer, derived from the plasma membrane of a host cell, that contains abundant viral glycoproteins. The processes of adsorption and release of enveloped viruses differ substantially from these processes for nonenveloped viruses. To illustrate lytic replication of enveloped viruses, we consider the rabies virus, whose nucleocapsid consists of a single-stranded RNA genome bound by multiple copies of a nucleocapsid protein. Like other lytic RNA viruses, the rabies virus is replicated in the cytoplasm and does not require host-cell nuclear enzymes. As shown in Figure 5-46, a rabies virion is adsorbed by endocytosis (see Chapter 14, Figures 14-1 and 14-2), and release of progeny virions occurs by budding from the host-cell plasma membrane. Budding virions are clearly visible in electron micrographs of infected cells, as illustrated in Figure 5-47. Many tens of thousands of progeny virions bud from an infected host cell before it dies.
Page 216
FIGURE 5-46Lytic replication cycle of an enveloped animal virus. The rabies virus is an enveloped virus with a single-stranded RNA genome. The structural components of this virus are depicted at the top. After a virion adsorbs to multiple copies of a specific host membrane protein (step 1), the cell engulfs it in an endosome (step 2). A cellular protein in the endosome membrane pumps H+ ions from the cytosol into the endosome interior. The resulting decrease in endosomal pH induces a conformational change in the viral glycoprotein, leading to fusion of the viral envelope with the endosomal lipid bilayer and release of the nucleocapsid into the cytosol (steps 3 and 4). Viral RNA polymerase uses ribonucleoside triphosphates in the cytosol to replicate the viral RNA genome (step 5) and to synthesize viral mRNAs (step 1). One of the viral mRNAs encodes the viral transmembrane glycoprotein, which is inserted into the membrane of the endoplasmic reticulum (ER) as it is synthesized on ER-bound ribosomes (step 7). Carbohydrate is added to the large folded domain inside the ER lumen and is modified as the membrane and the associated glycoproteins pass through the Golgi apparatus (step 8). Vesicles with mature glycoprotein fuse with the host-cell plasma membrane, depositing viral glycoprotein on the cell surface with the large receptor-binding domain outside the cell (step 9). Meanwhile, other viral mRNAs are translated on host-cell ribosomes into nucleocapsid protein, matrix protein, and viral RNA polymerase (step 10). These proteins are assembled with replicated viral genomic RNA (bright red) into progeny nucleocapsids (step 11), which then associate with the cytosolic domain of viral transmembrane glycoproteins in the host-cell plasma membrane (step 12). The plasma membrane is folded around the nucleocapsid, forming a “bud” that is eventually released (step 13).
[Thomas Deerinck, NCMIR/Science Source.]
EXPERIMENTAL FIGURE 5-47Release of progeny virions by budding. Progeny virions of enveloped viruses are released by budding from infected cells. In this transmission electron micrograph of a cell infected with measles virus, virion buds are clearly visible protruding from the cell surface. Measles virus is an enveloped RNA virus with a helical nucleocapsid, like rabies virus, and replicates as illustrated in Figure 5-46.