Different Types of RNA Exhibit Various Conformations Related to Their Functions
The primary structure of RNA is generally similar to that of DNA, with two exceptions: the sugar component of RNA, ribose, has a hydroxyl group at the 2′ position (see Figure 2-16b), and thymine in DNA is replaced by uracil in RNA. The presence of thymine rather than uracil in DNA is important to the long-term stability of DNA because of its function in DNA repair (see Section 5.6). As noted earlier, the hydroxyl group on the 2′ carbon of ribose makes RNA more chemically labile than DNA. As a result of this lability, RNA is cleaved into mononucleotides by an alkaline solution (see Figure 5-6), whereas DNA is not. The 2′-hydroxyl group of RNA also provides a chemically reactive group that takes part in RNA-mediated catalysis. Like DNA, RNA is a long polynucleotide that can be double-stranded or single-stranded, linear or circular. It can also participate in a hybrid helix composed of one RNA strand and one DNA strand. As mentioned above, RNA-RNA and RNA-DNA double helices have a compact conformation like the A form of DNA (see Figure 5-4b).
Page 175
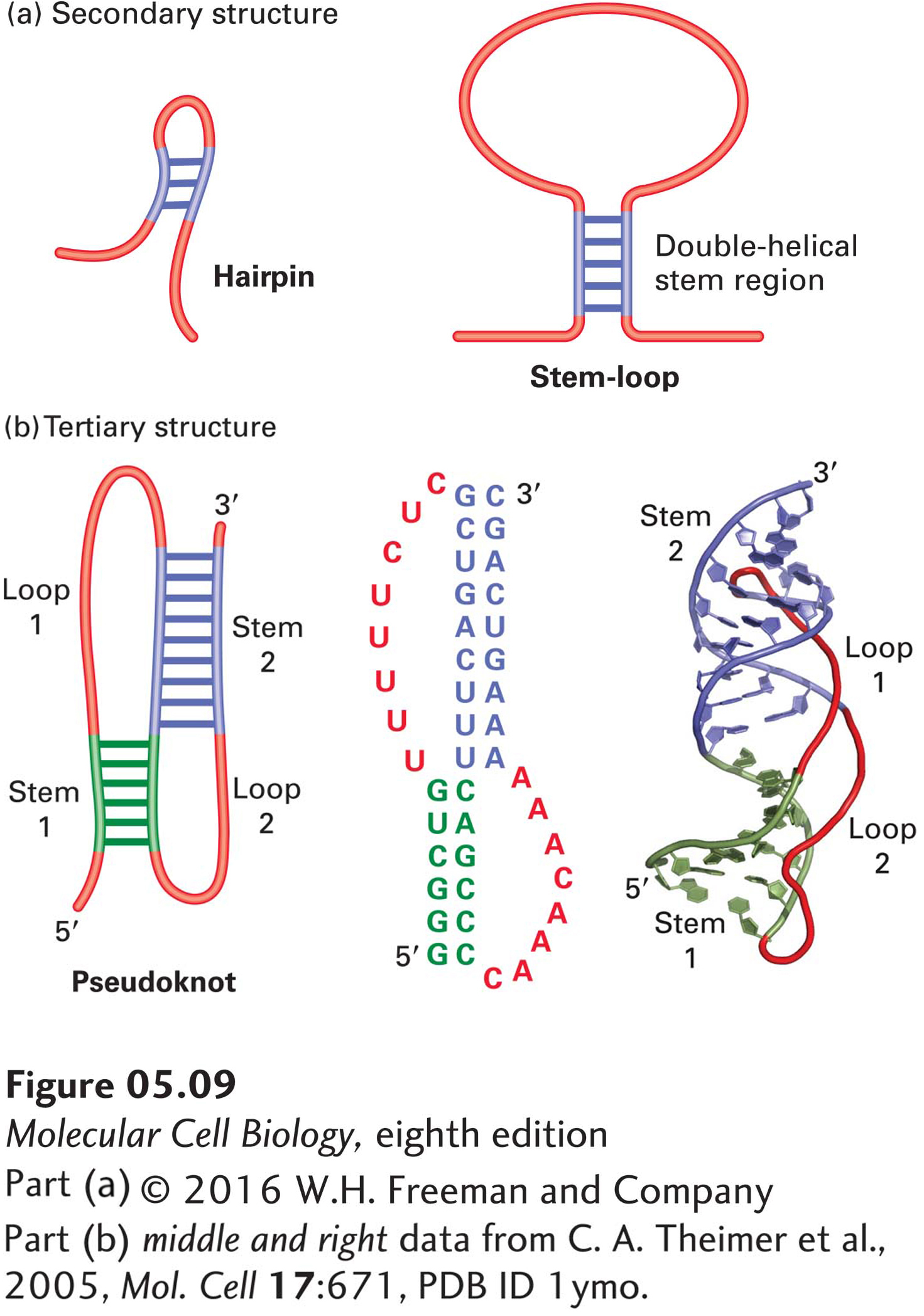
Unlike DNA, which exists primarily as a very long double helix, most cellular RNAs are single-stranded, and they exhibit a variety of conformations (Figure 5-9). Differences in the sizes and conformations of the various types of RNA permit them to carry out specific functions in a cell. The simplest secondary structures in single-stranded RNAs are formed by pairing of complementary bases. “Hairpins” are formed by pairing of bases within about five to ten nucleotides of each other, and “stem-loops” by pairing of bases that are separated by eleven to several hundred nucleotides. These simple folds can cooperate to form more complicated tertiary structures, one of which is termed a “pseudoknot.”
[Part (b) middle and right data from C. A. Theimer et al., 2005, Mol. Cell17:671, PDB ID 1ymo.]
FIGURE 5-9RNA secondary and tertiary structures. (a) Hairpins, stem-loops, and other secondary structures can form by base pairing between distant complementary segments of an RNA molecule. In stem-loops, the single-stranded loop between the base-paired helical stem may be hundreds or even thousands of nucleotides long, whereas in hairpins, the short turn may contain as few as four nucleotides. (b) Pseudoknots, one type of RNA tertiary structure, are formed by interaction of loops through base pairing between complementary bases. The structure shown forms the core domain of the human telomerase RNA. Left: Secondary-structure diagram with base-paired nucleotides in green and blue and single-stranded regions in red. Middle: Sequence of the telomerase RNA core domain, colored to correspond to the secondary-structure diagram at the left. Right: Diagram of the telomerase core domain structure determined by 2D-NMR, showing paired bases only and a tube for the sugar phosphate backbone, colored to correspond to the diagrams at left.
[Part (b) middle and right data from C. A. Theimer et al., 2005, Mol. Cell17:671, PDB ID 1ymo.]
As discussed in detail later, tRNA molecules adopt a well-defined three-dimensional architecture in solution that is crucial to protein synthesis. Larger rRNA molecules also have locally well-defined three-dimensional structures with more flexible linkers in between. Secondary and tertiary structures have also been recognized in mRNA, particularly near the ends of molecules. Clearly, then, RNA molecules are like proteins in that they have structured domains connected by less structured, flexible stretches.
The folded domains of RNA molecules not only are structurally analogous to the α helices and β strands found in proteins, but in some cases also have catalytic capacities. Such catalytic RNAs are called ribozymes. Although ribozymes are usually associated with proteins that stabilize the ribozyme structure, it is the RNA that acts as a catalyst. Some ribozymes can catalyze splicing, a remarkable process in which an internal RNA sequence is cut and removed and the two resulting chains are then ligated. This process occurs during the formation of the majority of functional mRNA molecules in multicellular eukaryotes, and it also occurs in single-celled eukaryotes such as yeasts, bacteria, and archaea. Remarkably, some RNAs carry out self-splicing, with the catalytic activity residing in the sequence that is removed. The mechanisms of splicing and self-splicing are discussed in detail in Chapter 10. As noted later in this chapter, rRNA plays a catalytic role in the formation of peptide bonds during protein synthesis.
In this chapter, we focus on the functions of mRNA, tRNA, and rRNA in gene expression. In later chapters, we will encounter other RNAs, often associated with proteins, that participate in other cell functions.