RNA Interference Causes Gene Inactivation by Destroying the Corresponding mRNA
The phenomenon known as RNA interference provides what may be the most straightforward method to inhibit the function of specific genes. This approach to disrupting genes is technically simpler than the methods described above. First observed in the roundworm C. elegans, RNA interference (RNAi) refers to the ability of double-stranded RNA to block expression of its corresponding single-stranded mRNA, but not that of mRNAs with a different sequence.
As described in Chapter 10, the phenomenon of RNAi rests on the general ability of eukaryotic cells to cleave double-stranded RNA into short (23-nt) double-stranded segments known as small inhibitory RNAs (siRNAs). The RNA endonuclease that catalyzes this reaction, known as Dicer, is found in all metazoans, but not in simpler eukaryotes such as yeast. The siRNA molecules, in turn, can cause cleavage of mRNA molecules of matching sequence, in a reaction catalyzed by a protein complex known as RISC. RISC mediates recognition and hybridization between one strand of the siRNA and its complementary sequence on the target mRNA; subsequently, specific nucleases in the RISC complex cleave the mRNA-siRNA hybrid. This model accounts for the specificity of RNAi, since it depends on base pairing, and for its potency in silencing gene function, since the complementary mRNA is permanently destroyed by nucleolytic degradation. The normal function of both Dicer and RISC is to allow for gene regulation by small endogenous RNA molecules known as micro-RNAs, or miRNAs.
Page 265
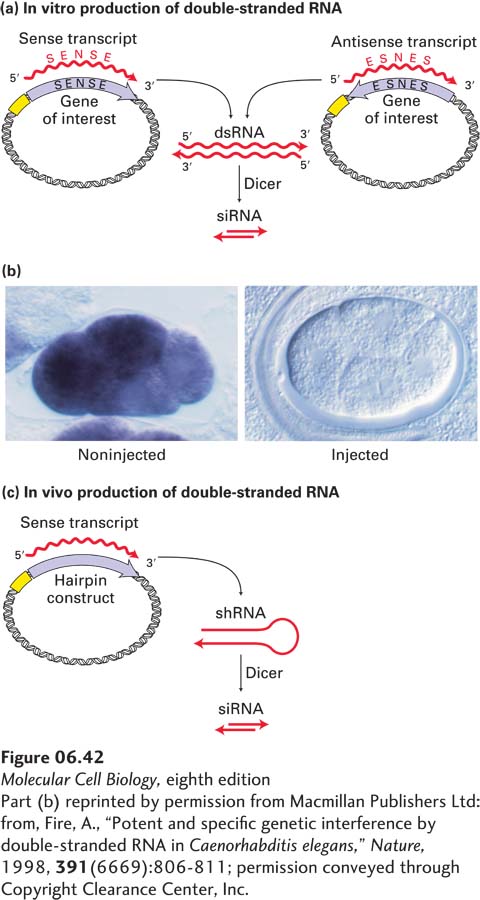
Researchers can exploit the micro-RNA pathway to intentionally silence a C. elegans gene of interest by using either of two general methods for generating siRNAs of defined sequence. In the first method, a double-stranded RNA corresponding to the target-gene sequence is produced by in vitro transcription of both sense and antisense copies of that sequence (Figure 6-42a). This dsRNA is then injected into the gonad of an adult worm, where it is converted to siRNA by Dicer in the developing embryos. In conjunction with the RISC complex, the siRNA molecules cause the corresponding mRNA molecules to be destroyed rapidly. The resulting worms display a phenotype similar to the one that would result from disruption of the target gene itself. In some cases, insertion of just a few molecules of a particular dsRNA into a cell is sufficient to inactivate many copies of the corresponding mRNA. Figure 6-42b illustrates the ability of an injected dsRNA to interfere with the production of the corresponding endogenous mRNA in C. elegans embryos. In this experiment, the mRNA levels in embryos were determined by in situ hybridization, as described earlier, using a fluorescently labeled probe.
[Part (b) reprinted by permission from Macmillan Publishers Ltd: from, Fire, A., “Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans,” Nature, 1998, 391(6669):806-811; permission conveyed through Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 6-42RNA interference (RNAi) can inhibit gene function in C. elegans and other organisms. (a) In vitro production of double-stranded RNA (dsRNA) for interference with a specific target gene. The coding sequence of the gene, derived from either a cDNA clone or a segment of genomic DNA, is placed in two orientations in a plasmid vector adjacent to a strong promoter. Transcription of both constructs in vitro using RNA polymerase and ribonucleoside triphosphates yields many RNA copies in both the sense orientation (identical to the mRNA sequence) and the complementary antisense orientation. Under suitable conditions, these complementary RNA molecules hybridize to form dsRNA. When the dsRNA is injected into cells, it is cleaved by Dicer into siRNAs. (b) Inhibition of mex3 RNA expression in C. elegans embryos by RNAi (see the text for the mechanism). Expression of mex3 RNA in embryos was assayed by in situ hybridization to a probe specific for this mRNA, linked to an enzyme that produces a colored (purple) product. (Left) Wild-type embryo. (Right) Embryo derived from a worm injected with double-stranded mex3 RNA. Each four-cell-stage embryo is ∼50 mm in length. (c) In vivo production of double-stranded RNA via an engineered plasmid introduced directly into cells. The synthetic gene construct is a tandem arrangement of both sense and antisense sequences of the target gene. When it is transcribed, double-stranded small hairpin RNA (shRNA) forms. The shRNA is cleaved by Dicer to form siRNA.
[Part (b) reprinted by permission from Macmillan Publishers Ltd: from, Fire, A., “Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans,” Nature, 1998, 391(6669):806-811; permission conveyed through Copyright Clearance Center, Inc.]
Page 266
The second method is to produce a specific double-stranded RNA in vivo. An efficient way to do this is to express a synthetic gene that is designed to contain tandem segments of both sense and antisense sequences corresponding to the target gene (Figure 6-42c). When this gene is transcribed, a double-stranded RNA “hairpin” structure forms, known as small hairpin RNA, or shRNA. The shRNA will then be cleaved by Dicer to form siRNA molecules. The lentiviral expression vectors are particularly useful for introducing synthetic genes for the expression of shRNA constructs into animal cells.
Both RNAi methods lend themselves to systematic studies in which researchers inactivate each of the known genes in an organism and observe what goes wrong. For example, in initial studies with C. elegans, RNA interference with 16,700 genes (about 86 percent of the genome) yielded 1722 visibly abnormal phenotypes. The genes whose functional inactivation causes particular abnormal phenotypes can be grouped into sets; each member of a set presumably controls the same signals or events. The regulatory relations between the genes in a set—for example, the genes that control muscle development—can then be worked out.
RNAi-mediated gene inactivation has been successful in many organisms, including Drosophila, many kinds of plants, zebrafish, the frog Xenopus, and mice, and these organisms are now the subjects of large-scale RNAi screens. For example, lentiviral vectors have been designed to inactivate by RNAi more than 10,000 different genes expressed in cultured mammalian cells. The functions of the inactivated genes can be inferred from defects in the growth or morphology of cell clones transfected with lentiviral vectors.