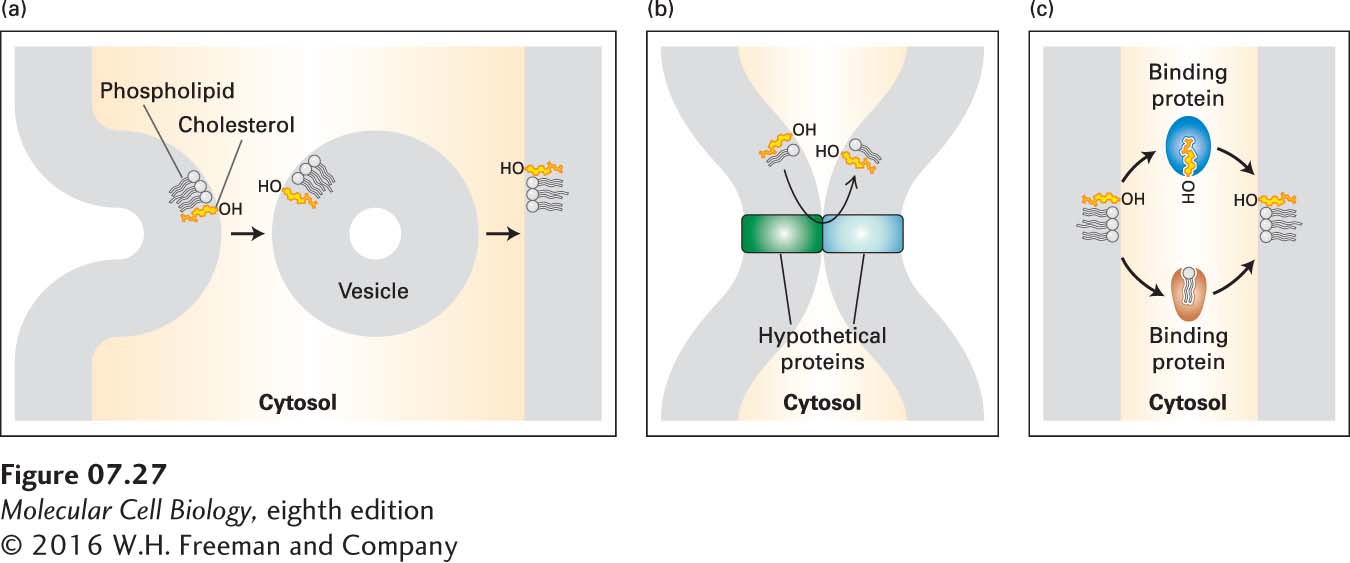
Cholesterol and Phospholipids Are Transported Between Organelles by Several Mechanisms
As already noted, the final steps in the synthesis of cholesterol and phospholipids take place primarily in the ER. Thus the plasma membrane and the membranes bounding other organelles must obtain these lipids by means of one or more intracellular transport processes. Membrane lipids can and do accompany both soluble and membrane proteins along the secretory pathway described in Chapter 14: membrane vesicles bud from the ER and fuse with membranes in the Golgi complex, and other membrane vesicles bud from the Golgi complex and fuse with the plasma membrane (Figure 7-27a). However, several lines of evidence suggest that there is substantial inter-organelle movement of cholesterol and phospholipids through other mechanisms. For example, chemical inhibitors of the classic secretory pathway and mutations that impede vesicular traffic along this pathway do not prevent cholesterol or phospholipid transport between membranes.
FIGURE 7-27Proposed mechanisms of transport of cholesterol and phospholipids between membranes. In mechanism (a), vesicles transfer lipids between membranes. In mechanism (b), lipid transfer is a consequence of direct contact between membranes that is mediated by membrane-embedded proteins. In mechanism (c), transfer is mediated by small, soluble lipid-transfer proteins. See F. R. Maxfield and D. Wustner, 2002, J. Clin. Invest.110:891.
Page 297
A second proposed mechanism of lipid movement entails direct protein-mediated contact of ER or ER-derived membranes with the membranes of other organelles (Figure 7-27b). In a third proposed mechanism, small lipid-transfer proteins facilitate the exchange of phospholipids or cholesterol between different membranes (Figure 7-27c). Although such lipid-transfer proteins have been identified in assays in vitro, their role in the intracellular movements of most phospholipids is not well defined. For instance, mice with a knockout mutation in the gene encoding the phosphatidylcholine-transfer protein appear to be normal in most respects, indicating that this protein is not essential for cellular phospholipid metabolism.
As noted earlier, the lipid compositions of different organelle membranes vary considerably (see Table 7-1). Some of these differences are due to different sites of synthesis. For example, a phospholipid called cardiolipin, which is localized to the mitochondrial membrane, is made only in mitochondria, and little of it is transferred to other organelles. Differential transport of lipids also plays a role in determining the lipid compositions of different cellular membranes. For instance, even though cholesterol is made in the ER, the cholesterol concentration (cholesterol-to-phospholipid molar ratio) is 1.5–13-fold higher in the plasma membrane than in other organelles (ER, Golgi, mitochondrion, lysosome). Although the mechanisms responsible for establishing and maintaining these differences are not well understood, we have seen that the distinctive lipid composition of each membrane has a major influence on its physical and biological properties.