Most Lipids and Many Proteins Are Laterally Mobile in Biomembranes
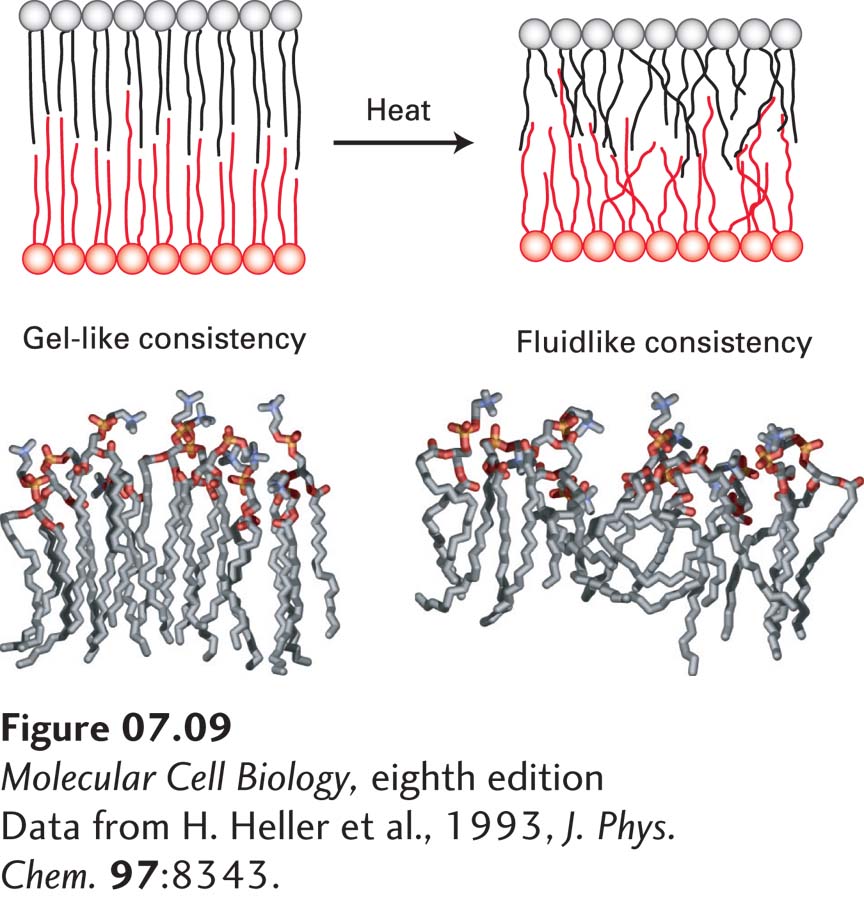
In the two-dimensional plane of a bilayer, thermal motion permits lipid molecules to rotate freely around their long axes and to diffuse laterally within each leaflet. Because such movements are lateral or rotational, the fatty acyl chains remain in the hydrophobic interior of the bilayer. In both natural and artificial membranes, a typical lipid molecule exchanges places with its neighbors in a leaflet about 107 times per second and diffuses several micrometers per second at 37 °C. These diffusion rates indicate that the bilayer is 100 times more viscous than water—about the same as the viscosity of olive oil. Even though lipids diffuse more slowly in the bilayer than in an aqueous solvent, a membrane lipid could diffuse the length of a typical bacterial cell (1 µm) in only 1 second and the length of an animal cell in about 20 seconds. When artificial pure phospholipid membranes are cooled below 37 °C, the lipids can undergo a phase transition from a liquid-like (fluid) state to a gel-like (semisolid) state, analogous to the liquid-solid transition when liquid water freezes (Figure 7-9). Below the phase-transition temperature, the rate of diffusion of the lipids drops precipitously. At usual physiological temperatures, the hydrophobic interior of natural membranes generally has a low viscosity and a fluidlike consistency, in contrast to the gel-like consistency observed at lower temperatures.
[Data from H. Heller et al., 1993, J. Phys. Chem.97:8343.]
FIGURE 7-9Gel-like and fluidlike forms of the phospholipid bilayer. (Top) Depiction of gel-to-fluid transition. Phospholipids with long saturated fatty acyl chains tend to assemble into a highly ordered, gel-like bilayer in which there is little overlap of the nonpolar tails in the two leaflets. Heat disorders the nonpolar tails and induces a transition from a gel to a fluid within a temperature range of only a few degrees. As the chains become disordered, the bilayer also decreases in thickness. (Bottom) Molecular models of phospholipid monolayers in gel-like and fluidlike states, as determined by molecular dynamics calculations.
[Data from H. Heller et al., 1993, J. Phys. Chem.97:8343.]
Page 279
In pure lipid bilayers (i.e., in the absence of protein), phospholipids and sphingolipids rotate and move laterally, but they do not spontaneously migrate, or flip-flop, from one leaflet to the other. The energetic barrier is too high; migration would require moving the polar head group from its aqueous environment through the hydrocarbon core of the bilayer to the aqueous solution on the other side. Special membrane proteins discussed in Chapter 11 are required to flip membrane lipids and other polar molecules from one leaflet to the other.
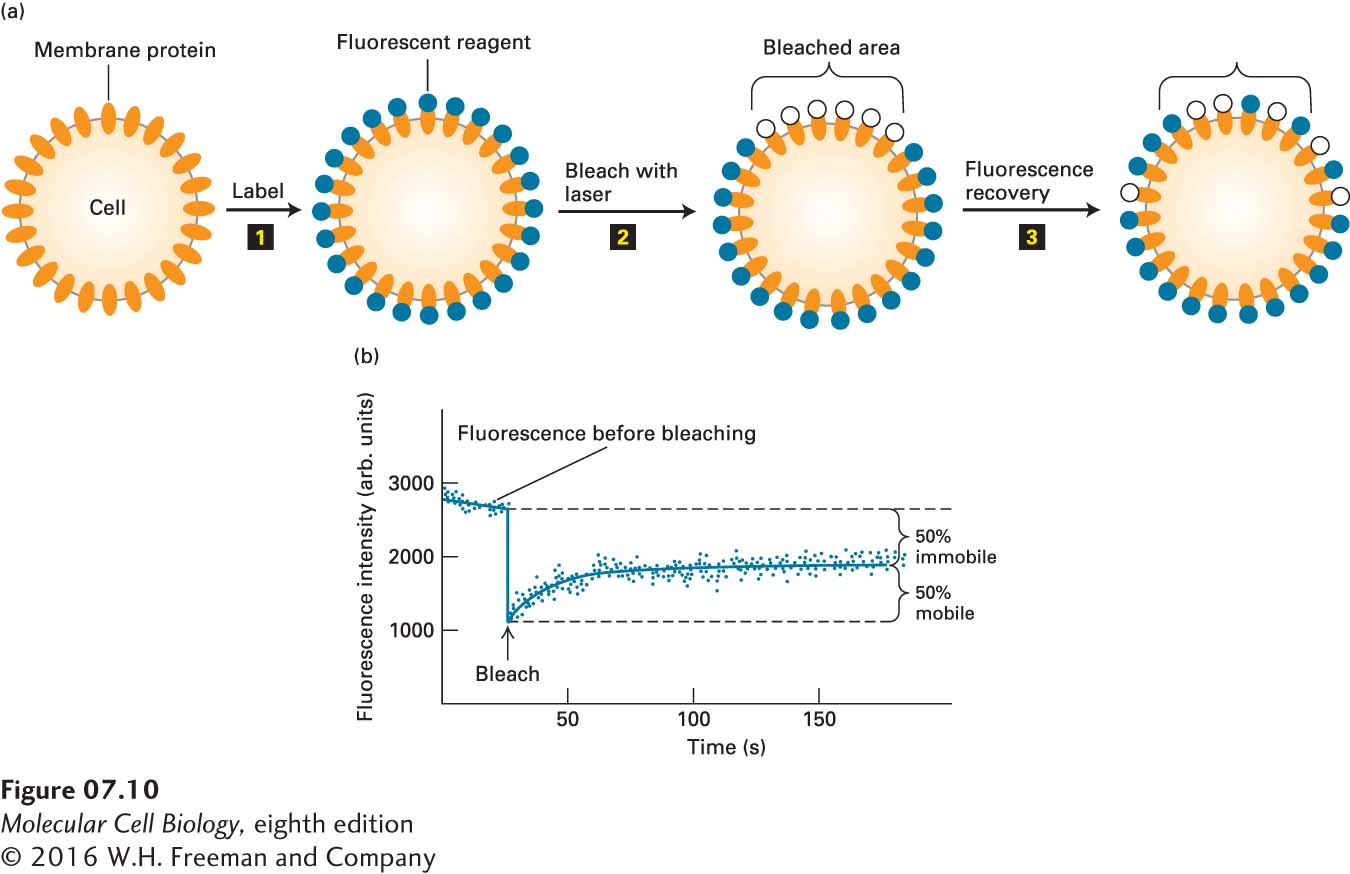
The lateral movements of specific plasma-membrane proteins and lipids can be quantified by a technique called fluorescence recovery after photobleaching (FRAP). Phospholipids containing a fluorescent substituent are used to monitor lipid movement. For proteins, a fragment of a monoclonal antibody that is specific for the exoplasmic domain of the desired protein and that has only a single antigen-binding site is tagged with a fluorescent dye. With this method, described in Figure 7-10, the rate at which membrane molecules move—the diffusion coefficient—can be determined, as well as the proportion of the molecules that are laterally mobile.
EXPERIMENTAL FIGURE 7-10Fluorescence recovery after photobleaching (FRAP) experiments can quantify the lateral movement of proteins and lipids within the plasma membrane. (a) Experimental protocol. Step 1: Cells are first labeled with a fluorescent reagent that binds uniformly to a specific membrane lipid or protein. Step 2: A laser light is then focused on a small area of the cell surface, irreversibly bleaching the bound reagent and thus reducing the fluorescence in the illuminated area. Step 3: In time, the fluorescence of the bleached patch increases as unbleached fluorescent surface molecules diffuse into it and bleached ones diffuse outward. The extent of recovery of fluorescence in the bleached patch is proportional to the fraction of labeled molecules that are mobile in the membrane. (b) Results of a FRAP experiment with human hepatoma cells treated with a fluorescent antibody specific for the asialoglycoprotein receptor protein. The finding that 50 percent of the fluorescence returned to the bleached area indicates that 50 percent of the receptor molecules in the illuminated membrane patch were mobile and 50 percent were immobile. Because the rate of fluorescence recovery is proportional to the rate at which labeled molecules move into the bleached region, the diffusion coefficient of a protein or lipid in the membrane can be calculated from such data. See Y. I. Henis et al., 1990, J. Cell Biol.111:1409.
The results of FRAP studies with fluorescence-labeled phospholipids have shown that in fibroblast plasma membranes, all the phospholipids are freely mobile over distances of about 0.5 µm, but most cannot diffuse over much longer distances. These findings suggest that protein-rich regions of the plasma membrane about 1 µm in diameter separate lipid-rich regions containing the bulk of the membrane phospholipids. Phospholipids are free to diffuse within such regions, but not from one lipid-rich region to an adjacent one. Furthermore, the rate of lateral diffusion of lipids in the plasma membrane is nearly an order of magnitude slower than in pure phospholipid bilayers: diffusion constants of 10−8 cm2 per second and 10−7 cm2 per second are characteristic of the plasma membrane and a pure phospholipid bilayer, respectively. This difference suggests that lipids may be tightly but not irreversibly bound to certain integral proteins in some membranes, as indeed has recently been demonstrated (see the discussion of annular phospholipids below).