Elongation Factors Regulate the Initial Stages of Transcription in the Promoter-Proximal Region
In metazoans, at most promoters, Pol II pauses after transcribing fewer than 100 nucleotides, due to the binding of a five-subunit protein called NELF (negative elongation factor). NELF binds to Pol II along with a two-subunit elongation factor called DSIF (DRB sensitivity-inducing factor, so named because an ATP analog called DRB inhibits further transcription elongation in its presence). The inhibition of elongation that results from NELF binding to Pol II is relieved when DSIF, NELF, and serine 2 of the Pol II CTD (Tyr-Ser-Pro-Thr-Ser-Pro-Ser) are phosphorylated by a protein kinase with two subunits, cyclin T–CDK9, also called P-TEFb, which associates with the Pol II-NELF-DSIF complex. The same elongation factors regulate transcription from CpG island promoters. These factors that regulate elongation in the promoter-proximal region provide a mechanism for controlling gene transcription in addition to the regulation of transcription initiation. This overall strategy for regulating transcription at both the initiation and elongation steps in the promoter-proximal region is similar to the regulation of the trp operon in E. coli (see Figure 9-7), although the molecular mechanisms involved are distinct.
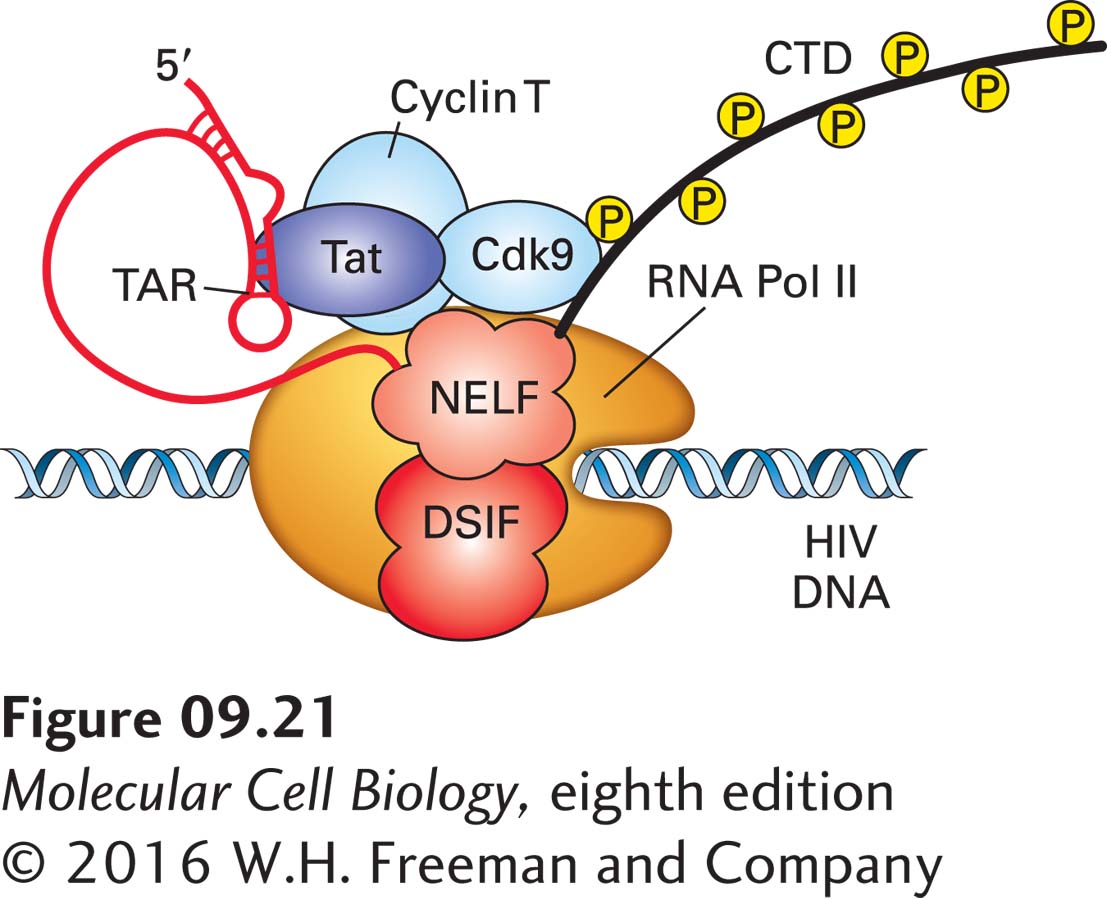
Transcription of HIV (human immunodeficiency virus), the cause of AIDS, is dependent on the activation of cyclin T–CDK9 by a small viral protein called Tat. Cells experimentally infected with tat− mutants produce short viral transcripts about 50 nucleotides long. In contrast, cells infected with wild-type HIV synthesize long viral transcripts that encompass the entire integrated proviral genome (see Figure 5-48 and Figure 8-13). Thus Tat functions as an antitermination factor, permitting RNA polymerase II to read through a transcriptional block. (Tat is initially made by rare transcripts that fail to terminate when the HIV promoter is transcribed at a high rate in “activated” T-lymphocytes; see Chapter 23.) Tat is a sequence-specific RNA-binding protein. It binds to the RNA copy of a sequence called TAR, which forms a stem-loop structure near the 5′ end of the HIV transcript (Figure 9-21). TAR also binds cyclin T, holding the cyclin T–CDK9 complex close to the polymerase, where it efficiently phosphorylates its substrates, resulting in transcription elongation. Chromatin immunoprecipitation assays done after treating cells with specific inhibitors of CDK9 indicate that the transcription of some 30 percent of mammalian genes is regulated by controlling the activity of cyclin T–CDK9 (P-TEFb), although this is probably done most frequently by sequence-specific DNA-binding transcription factors rather than by an RNA-binding protein, as in the case of HIV Tat.
FIGURE 9-21Model of antitermination complex composed of HIV Tat protein and several cellular proteins. The TAR element in the HIV transcript contains sequences recognized by Tat and the cellular protein cyclin T. Cyclin T activates and helps position the protein kinase CDK9 near its substrates, the CTD of RNA polymerase II, NELF, and DSIF. CTD phosphorylation at serine 2 of the Pol II CTD heptad repeat is required for transcription elongation. Cellular proteins DSIF and the NELF complex are also involved in regulating Pol II elongation, as discussed in the text. See T. Wada et al., 1998, Gene Dev.12:343; Y. Yamaguchi et al., 1999, Cell97:451; and T. Yamada et al., 2006, Mol. Cell21:227.