Formation of Heterochromatin Silences Gene Expression at Telomeres, near Centromeres, and in Other Regions
For many years it has been clear that inactive genes in eukaryotic cells are often associated with heterochromatin, regions of chromatin that are more highly condensed and stain more darkly with DNA dyes than euchromatin, in which most transcribed genes are located (see Figure 8-28a). Regions of chromosomes near the centromeres and telomeres, as well as additional specific regions that vary in different cell types, are organized into heterochromatin. The DNA in heterochromatin is less accessible to externally added proteins than is DNA in euchromatin and consequently is often referred to as “closed” chromatin. For instance, in an experiment described in Chapter 8, the DNA of inactive genes was found to be far more resistant to digestion by DNase I than the DNA of transcribed genes (see Figure 8-27).
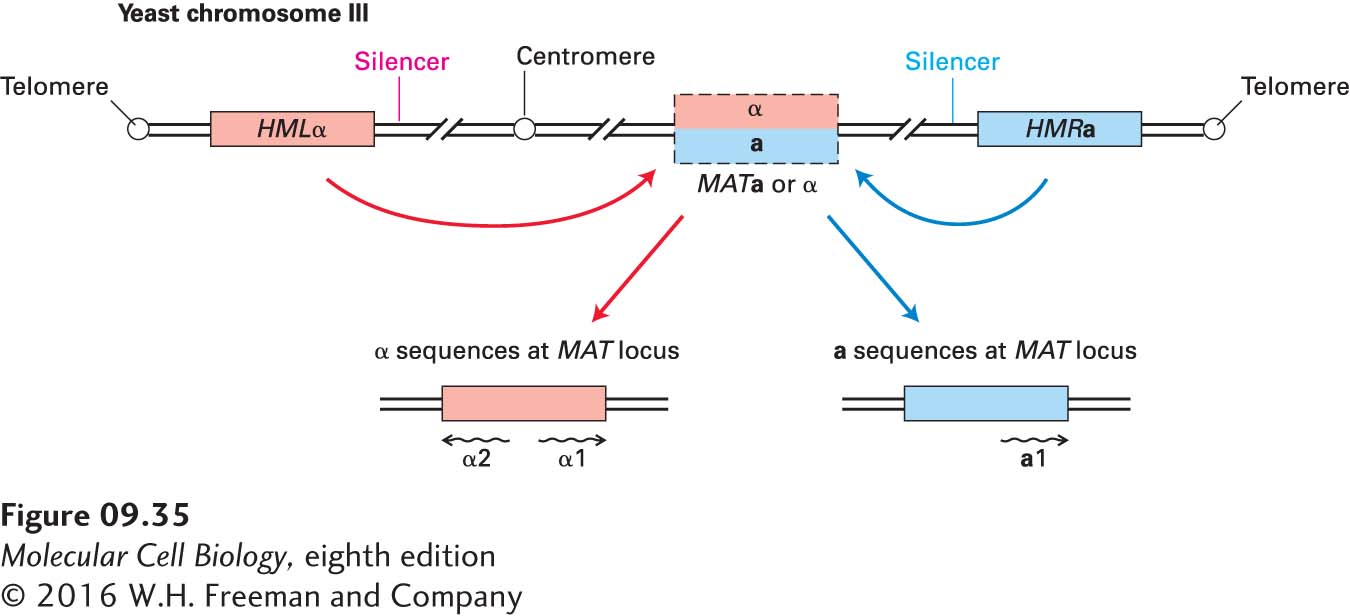
Study of DNA regions in S. cerevisiae that behave like the heterochromatin of higher eukaryotes provided early insight into the chromatin-mediated repression of transcription. This yeast can grow either as haploid or diploid cells. Haploid cells exhibit one of two possible mating types, called a and α. Cells of different mating type can “mate,” or fuse, to generate a diploid cell (see Figure 1-23). When a haploid cell divides by budding, the larger “mother” cell switches its mating type. Genetic and molecular analyses have revealed that three genetic loci on yeast chromosome III control the mating type of yeast cells (Figure 9-35). The central mating-type locus, termed MAT—the only one of the three that is actively transcribed—encodes transcription factors (a1, or α1 and α2) that regulate genes that determine the mating type. In any one cell, either an a or α DNA sequence is located at the MAT. The two additional loci, termed HML and HMR, near the left and right telomere, respectively, contain “silent” (nontranscribed) copies of the a or α genes. These sequences are transferred alternately from HMLα or HMRa into the MAT locus by a type of nonreciprocal recombination between homologous sequences during cell division. When the MAT locus contains the DNA sequence from HMLα, the cells behave as α cells. When the MAT locus contains the DNA sequence from HMRa, the cells behave like a cells.
FIGURE 9-35Arrangement of mating-type loci on chromosome III in the yeast S. cerevisiae. Silent (unexpressed) mating-type genes (either a or α) are located at the HML locus. The opposite mating-type gene is present at the silent HMR locus. When the α or a sequences are present at the MAT locus, they can be transcribed into mRNAs whose encoded proteins specify the mating-type phenotype of the cell. The silencer sequences near HML and HMR bind proteins that are critical for repression of these silent loci. Haploid cells can switch mating types in a process that transfers the DNA sequence from HML or HMR to the transcriptionally active MAT locus.
Page 391
Our interest here is in how transcription of the silent mating-type genes at HML and HMR is repressed. If these genes are expressed, as they are in yeast mutants with defects in the repressing mechanism, both a and α proteins are expressed, causing the cells to behave like diploid cells, which cannot mate. The promoters and UASs controlling transcription of the a and α genes lie near the center of the DNA sequence that is transferred and are identical whether the sequences are at the MAT locus or at one of the silent loci. This arrangement indicates that the function of the transcription factors that interact with these sequences must somehow be blocked at HML and HMR, but not at the MAT locus. This repression of the silent loci depends on silencer sequences located next to the region of transferred DNA at HML and HMR (see Figure 9-35). If the silencer is deleted, the adjacent locus is transcribed. Remarkably, any gene placed near the yeast mating-type silencer sequence by recombinant DNA techniques is repressed, or “silenced,” even a tRNA gene transcribed by RNA polymerase III, which uses a different set of general transcription factors than RNA polymerase II uses, as discussed later.
Several lines of evidence indicate that repression of the HML and HMR loci results from a condensed chromatin structure that sterically blocks transcription factors from interacting with the DNA. In one telling experiment, the gene encoding an E. coli enzyme that methylates adenine residues in the sequence GATC was introduced into yeast cells under the control of a yeast promoter so that the enzyme was expressed. Researchers found that GATC sequences within the MAT locus and most other regions of the genome in these cells were methylated, but not those within the HML and HMR loci. These results indicate that the DNA of the silent loci is inaccessible to the E. coli methylase, and presumably to proteins in general, including transcription factors and RNA polymerase. Similar experiments conducted with various yeast histone mutants indicated that specific interactions involving the histone tails of H3 and H4 are required for formation of a fully repressed chromatin structure. Other studies have shown that the telomeres of every yeast chromosome also behave like silencer sequences. For instance, when a gene is placed within a few kilobases of any yeast telomere, its expression is repressed. In addition, this repression is relieved by the same mutations in the H3 and H4 histone tails that interfere with repression at the silent mating-type loci.
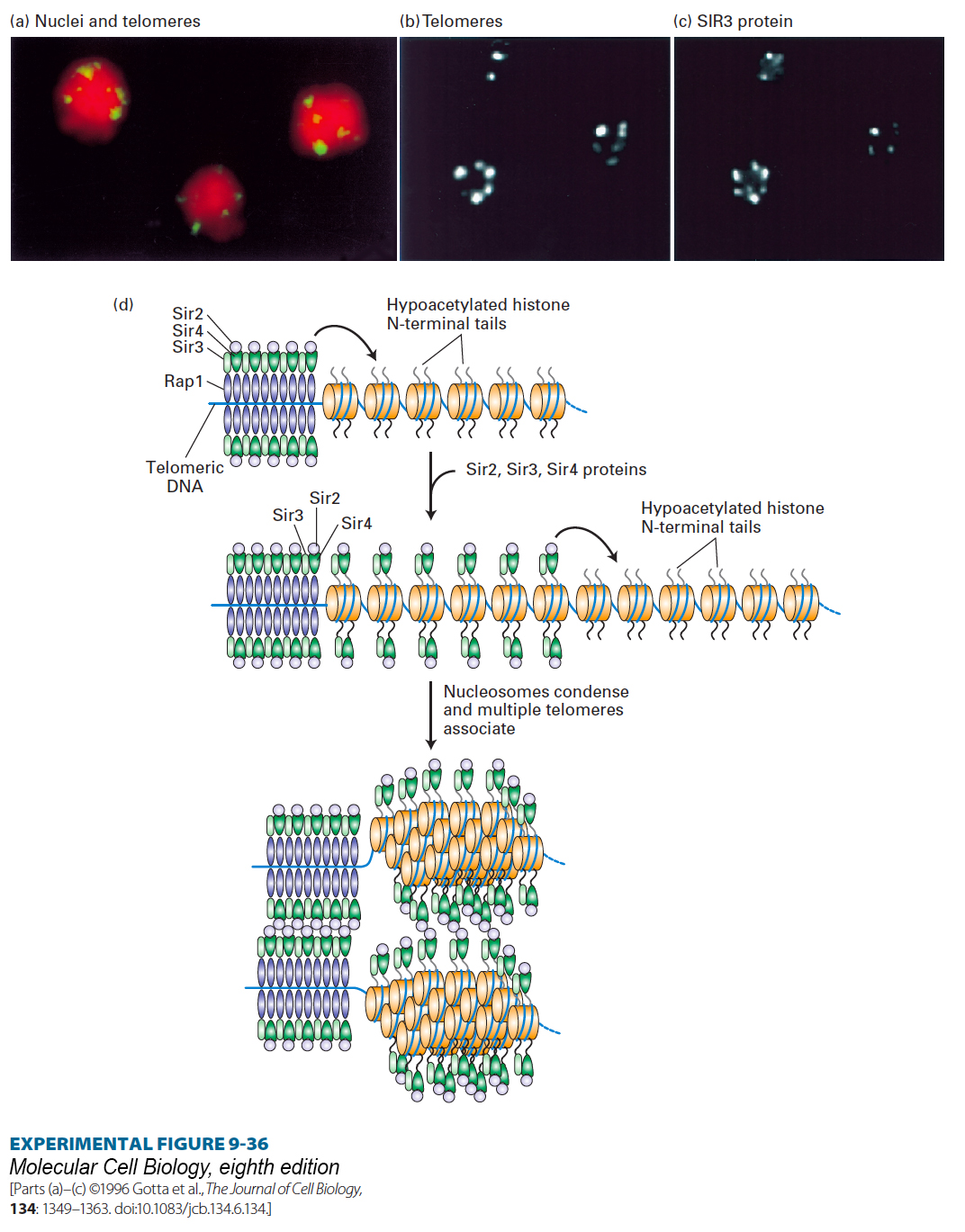
Genetic studies led to identification of several proteins, RAP1 and three SIR proteins, that are required for repression of the silent mating-type loci and the telomeres in yeast. RAP1 was found to bind within the DNA silencer sequences associated with HML and HMR and to a sequence that is repeated multiple times at each yeast-chromosome telomere. Further biochemical studies showed that the SIR2 protein is a histone deacetylase; it removes acetyl groups on lysines of the histone tails. Furthermore, the RAP1 and SIR2, 3, and 4 proteins bind to one another, and SIR3 and SIR4 bind to the N-terminal tails of histones H3 and H4, which are maintained in a largely nonacetylated state by the deacetylase activity of SIR2. A series of experiments using fluorescence confocal microscopy on yeast cells either stained with fluorescent-labeled antibody to any one of the SIR proteins or RAP1 or hybridized to a labeled telomere-specific DNA probe revealed that these proteins form large, condensed telomeric nucleoprotein structures resembling the heterochromatin found in higher eukaryotes (Figure 9-36a, b, c).
EXPERIMENTAL FIGURE 9-36Antibody and DNA probes colocalize SIR3 protein with telomeric heterochromatin in yeast nuclei. (a) Confocal micrograph 0.3 mm thick through three diploid yeast cells, each containing 68 telomeres. Telomeres were labeled by hybridization to a fluorescent telomere-specific probe (yellow). DNA was stained red to reveal the nuclei. The 68 telomeres coalesce into a much smaller number of regions near the nuclear periphery. (b, c) Confocal micrographs of yeast cells labeled with a telomere-specific hybridization probe (b) and a fluorescent-labeled antibody specific for SIR3 (c). Note that SIR3 is localized in the repressed telomeric heterochromatin. Similar experiments with RAP1, SIR2, and SIR4 have shown that these proteins also colocalize with the repressed telomeric heterochromatin. (d) Schematic model of the silencing mechanism at yeast telomeres. (Top left) Multiple copies of RAP1 bind to a simple repeated sequence at each telomere region that lacks nucleosomes. SIR3 and SIR4 bind to RAP1, and SIR2 binds to SIR4. SIR2 is a histone deacetylase that deacetylates the tails on the histones neighboring the repeated RAP1-binding site. (Middle) The hypoacetylated histone tails are also binding sites for SIR3 and SIR4, which in turn bind additional SIR2, deacetylating neighboring histones. Repetition of this process results in spreading of the region of hypoacetylated histones with associated SIR2, SIR3, and SIR4. (Bottom) Interactions between complexes of SIR2, SIR3, and SIR4 cause the chromatin to condense and several telomeres to associate, as shown in a–c. The higher-order chromatin structure generated sterically blocks other proteins from interacting with the underlying DNA. See M. Grunstein, 1997, Curr. Opin. Cell Biol. 9:383.
Figure 9-36d depicts a model for the chromatin-mediated silencing at yeast telomeres based on these and other studies. Formation of heterochromatin at telomeres is nucleated by multiple RAP1 protein molecules bound to repeated sequences in a nucleosome-free region at the extreme end of a telomere. A network of protein-protein interactions involving telomere-bound RAP1, three SIR proteins (2, 3, and 4), and hypoacetylated histones H3 and H4 creates a higher-order nucleoprotein complex that includes several telomeres and in which the DNA is largely inaccessible to external proteins. One additional protein, SIR1, is also required for silencing of the mating-type loci. It binds to the silencer regions associated with HML and HMR together with RAP1 and other proteins to initiate assembly of a similar multiprotein silencing complex that encompasses HML and HMR.
An important feature of this model is the dependence of repression on hypoacetylation of the histone tails. This dependence was demonstrated in experiments with yeast mutants expressing histones in which lysines in histone N-termini were replaced with arginines, glutamines, or glycines. Arginine is positively charged, like lysine, but cannot be acetylated. Glutamine, on the other hand, is neutral and simulates the neutral charge of acetylated lysine, and glycine, with no side chain, also mimics the absence of a positively charged lysine. Repression at telomeres and at the silent mating-type loci was defective in the mutants with glutamine and glycine substitutions for lysine in the H3 or H4 histone tails, but not in the mutants with arginine substitutions. Further, acetylation of H3 and H4 lysines interferes with binding by Sir3 and Sir4 and consequently prevents repression at the silent loci and telomeres. Finally, chromatin immunoprecipitation experiments (see Figure 9-18a) using antibodies specific for acetylated lysines at particular positions in the histone N-terminal tails (see Figure 8-26a) confirmed that histones in repressed regions near telomeres and at the silent mating loci are hypoacetylated, but become hyperacetylated in sir mutants when genes in these regions are derepressed.