Methylation of Specific Histone Lysines Is Linked to Epigenetic Mechanisms of Gene Repression
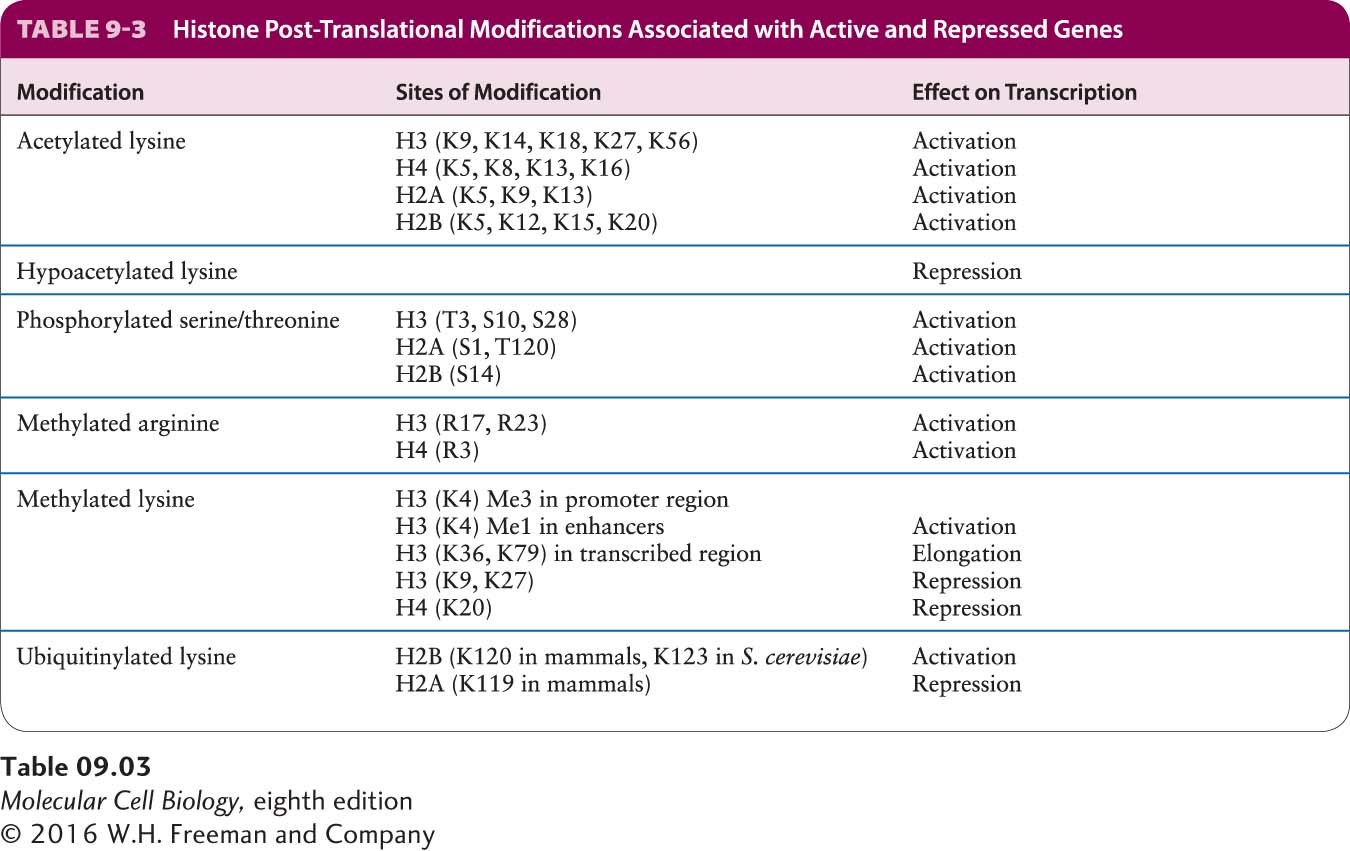
Figure 8-26b summarized the different types of post-translational modifications that are found on histones, including acetylation of lysines and methylation of lysines on the nitrogen atom of the terminal ε-amino group of the lysine side chain (see Figure 2-14). Lysines can be modified by the addition of one, two, or three methyl groups to this terminal nitrogen atom, generating mono-, di-, and trimethylated lysine, all of which carry a single positive charge.
The acetylation state at a specific histone lysine on a particular nucleosome results from a dynamic equilibrium between acetylation and deacetylation by histone acetylases and histone deacetylases, respectively. Acetylation of histones in a localized region of chromatin predominates when local DNA-bound activators transiently bind histone acetylase complexes. Deacetylation predominates when repressors transiently bind histone deacetylase complexes. Pulse-chase radiolabeling experiments have shown that acetyl groups on histone lysines turn over rapidly through the sequential actions of histone acetylases and histone deacetylases. In contrast, methyl groups on histones are much more stable. Histone lysine methyl groups can be removed by histone lysine demethylases. But the resulting turnover of histone lysine methyl groups is much slower than the turnover of histone lysine acetyl groups, which makes methylation the more appropriate post-translational modification for propagating epigenetic information.
Several other post-translational modifications of histones have been characterized (see Figure 8-26b). These modifications all have the potential to positively or negatively regulate the binding of proteins that interact with the chromatin fiber to regulate transcription as well as other processes, such as chromosome folding into the highly condensed structures that form during mitosis (see Figures 8-35 and 8-36). A picture of chromatin has emerged in which histone tails extending as random coils from the chromatin fiber are post-translationally modified to generate one of many possible combinations of modifications that regulate transcription and other processes by regulating the binding of a large number of different protein complexes. This control of the interactions of proteins with specific regions of chromatin that results from the combined influences of various post-translational modifications of histones has been called a histone code. Some of these modifications, such as histone lysine acetylation, are rapidly reversible, whereas others, such as histone lysine methylation, can be templated through chromatin replication, generating epigenetic inheritance in addition to inheritance of DNA sequence. Table 9-3 summarizes the influence that post-translational modifications of specific histone amino acid residues usually have on transcription.
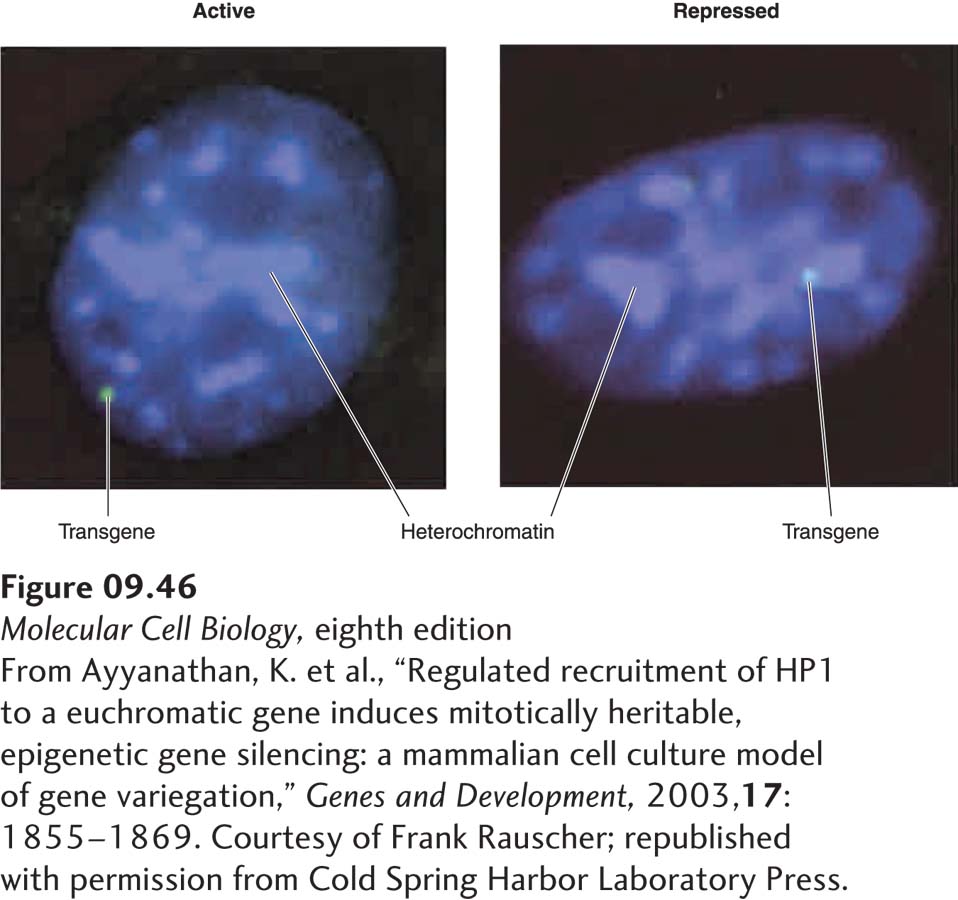
Histone H3 Lysine 9 Methylation in Heterochromatin In most eukaryotes, some co-repressor complexes contain histone methyl transferase subunits that methylate histone H3 at lysine 9, generating di- and trimethyl lysines. These methylated lysines are binding sites for isoforms of HP1 protein that function in the condensation of heterochromatin, as discussed in Chapter 8 (see Figure 8-29). For example, the KAP1 co-repressor complex functions with a class of more than 200 zinc-finger transcription factors encoded in the human genome. This co-repressor complex includes an H3 lysine 9 methyl transferase that methylates nucleosomes over the promoter regions of repressed genes, leading to HP1 binding and repression of transcription. An integrated transgene in cultured mouse fibroblasts that was repressed through the action of the KAP1 co-repressor was associated with heterochromatin in most cells, whereas the active form of the same transgene was associated with euchromatin (Figure 9-46). Chromatin immunoprecipitation assays (see Figure 9-18) showed that the repressed gene was associated with histone H3 methylated at lysine 9 and with HP1, whereas the active gene was not.
[From Ayyanathan, K. et al., “Regulated recruitment of HP1 to a euchromatic gene induces mitotically heritable, epigenetic gene silencing: a mammalian cell culture model of gene variegation,” Genes and Development, 2003, 17:1855–1869. Courtesy of Frank Rauscher; republished with permission from Cold Spring Harbor Laboratory Press.]
FIGURE 9-46Association of a repressed transgene with heterochromatin. Mouse fibroblasts were stably transformed with a transgene that contained binding sites for an engineered repressor. The repressor was a fusion between a DNA-binding domain, a repression domain that interacts with the KAP1 co-repressor complex, and the ligand-binding domain of a nuclear receptor that allows the nuclear import of the fusion protein to be controlled experimentally (see Figure 9-45). DNA was stained blue with the dye DAPI. Brighter-staining regions are regions of heterochromatin, where the DNA concentration is higher than in euchromatin. The transgene was detected by hybridization of a fluorescently labeled complementary probe (green). When the recombinant repressor was retained in the cytoplasm, the transgene was transcribed (left) and was associated with euchromatin in most cells. When hormone was added so that the recombinant repressor entered the nucleus, the transgene was repressed (right) and associated with heterochromatin. Chromatin immunoprecipitation assays (see Figure 9-18) showed that the repressed gene was associated with histone H3 methylated at lysine 9 and HP1, whereas the active gene was not.
[From Ayyanathan, K. et al., “Regulated recruitment of HP1 to a euchromatic gene induces mitotically heritable, epigenetic gene silencing: a mammalian cell culture model of gene variegation,” Genes and Development, 2003, 17:1855–1869. Courtesy of Frank Rauscher; republished with permission from Cold Spring Harbor Laboratory Press.]
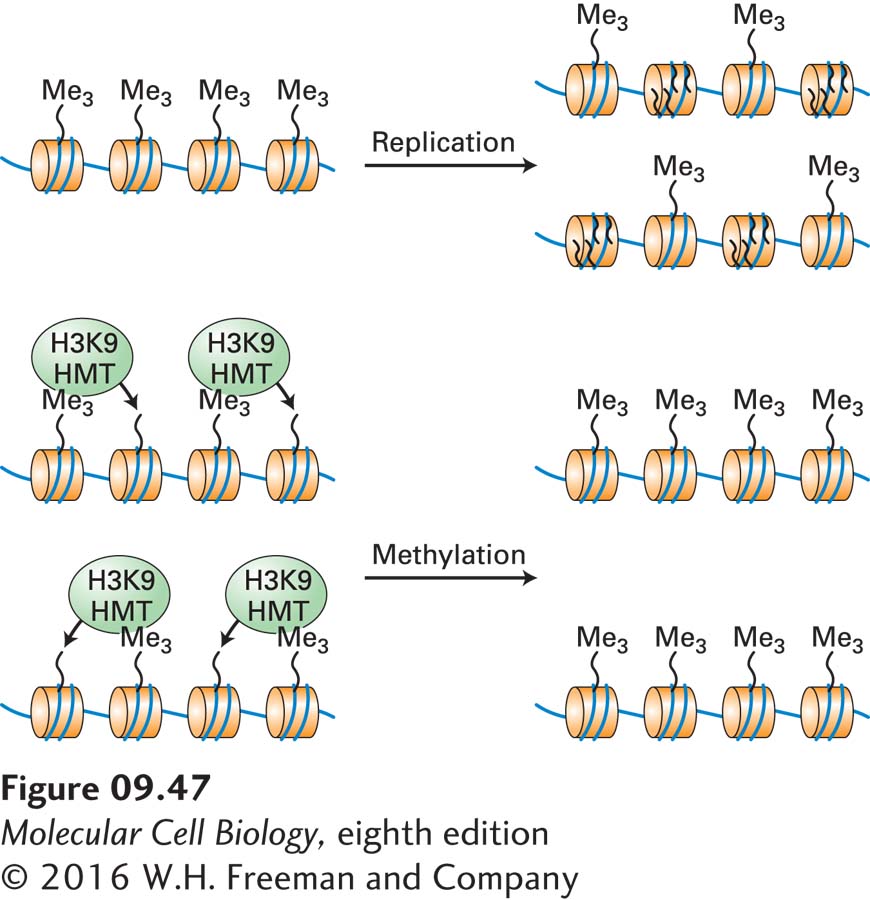
Importantly, H3 lysine 9 methylation is maintained following chromosome replication by the mechanism diagrammed in Figure 9-47. When a methylated region of DNA is replicated in S phase, the histone octomers associated with the parent DNA are randomly distributed to the daughter DNA molecules. New histone octamers that are not methylated on lysine 9 also associate randomly with the new daughter chromosomes, but since the parent histone octomers are associated with both daughter chromosomes, approximately half of the daughter chromosomes’ nucleosomes are methylated on lysine 9. Association of histone H3 lysine methyl transferases (directly or indirectly) with the parent methylated nucleosomes leads to methylation of the newly assembled histone octamers. Repetition of this process with each cell division results in maintenance of H3 lysine 9 methylation of this region of the chromosome.
FIGURE 9-47Maintenance of histone H3 lysine 9 methylation during chromosome replication. When chromosomal DNA is replicated, the parent histones randomly associate with the two daughter chromosomes, while unmethylated histones synthesized during S phase are assembled into other nucleosomes in those same daughter chromosomes. Association of histone H3 lysine 9 methyl transferases (H3K9 HMT) with parent nucleosomes bearing the histone 3 lysine 9 di- or trimethylation mark methylates the newly added unmodified nucleosomes. Consequently, histone H3 lysine 9 methylation marks are maintained during repeated cell divisions unless they are specifically removed by a histone demethylase.