Epigenetic Control by Polycomb and Trithorax Complexes
Another type of epigenetic mark that is essential for repression of genes in specific cell types in multicellular animals and plants involves a set of proteins known collectively as Polycomb proteins and a counteracting set of proteins known as Trithorax proteins. These names were derived from the phenotypes of mutations in the genes encoding these proteins in Drosophila, in which they were first discovered. The Polycomb repression mechanism is essential for maintaining the repression of genes in specific types of cells, and in all the subsequent cells that develop from them, throughout the life of an organism. Important genes regulated by Polycomb proteins include the Hox genes, which encode master regulatory transcription factors. Different combinations of Hox transcription factors help to direct the development of specific tissues and organs in a developing embryo. Early in embryogenesis, expression of Hox genes is controlled by typical activator and repressor proteins. However, the expression of these activators and repressors stops at an early point in embryogenesis. Correct expression of the Hox genes in the descendants of the early embryonic cells is then maintained throughout the remainder of embryogenesis and on into adult life by the Polycomb proteins, which maintain the repression of specific Hox genes. Trithorax proteins perform the opposite function, maintaining the expression of the Hox genes that were expressed in a specific cell early in embryogenesis in all the subsequent descendants of that cell. Polycomb and Trithorax proteins control thousands of genes, including genes that regulate cell growth and division (i.e., the cell cycle, as discussed in Chapter 19). Polycomb and Trithorax genes are often mutated in cancer cells, contributing importantly to the abnormal properties of these cells (see Chapter 24).
Page 407
Remarkably, virtually all cells in the developing embryo and adult express a similar set of Polycomb and Trithorax proteins, and all cells contain the same set of Hox genes. Yet only the Hox genes in cells where they were initially repressed in early embryogenesis remain repressed, even though the same Hox genes in other cells remain active in the presence of the same Polycomb proteins. Consequently, as in the case of the yeast silent mating-type loci, the expression of Hox genes is regulated by a process that involves more than specific DNA sequences interacting with proteins that diffuse through the nucleoplasm.
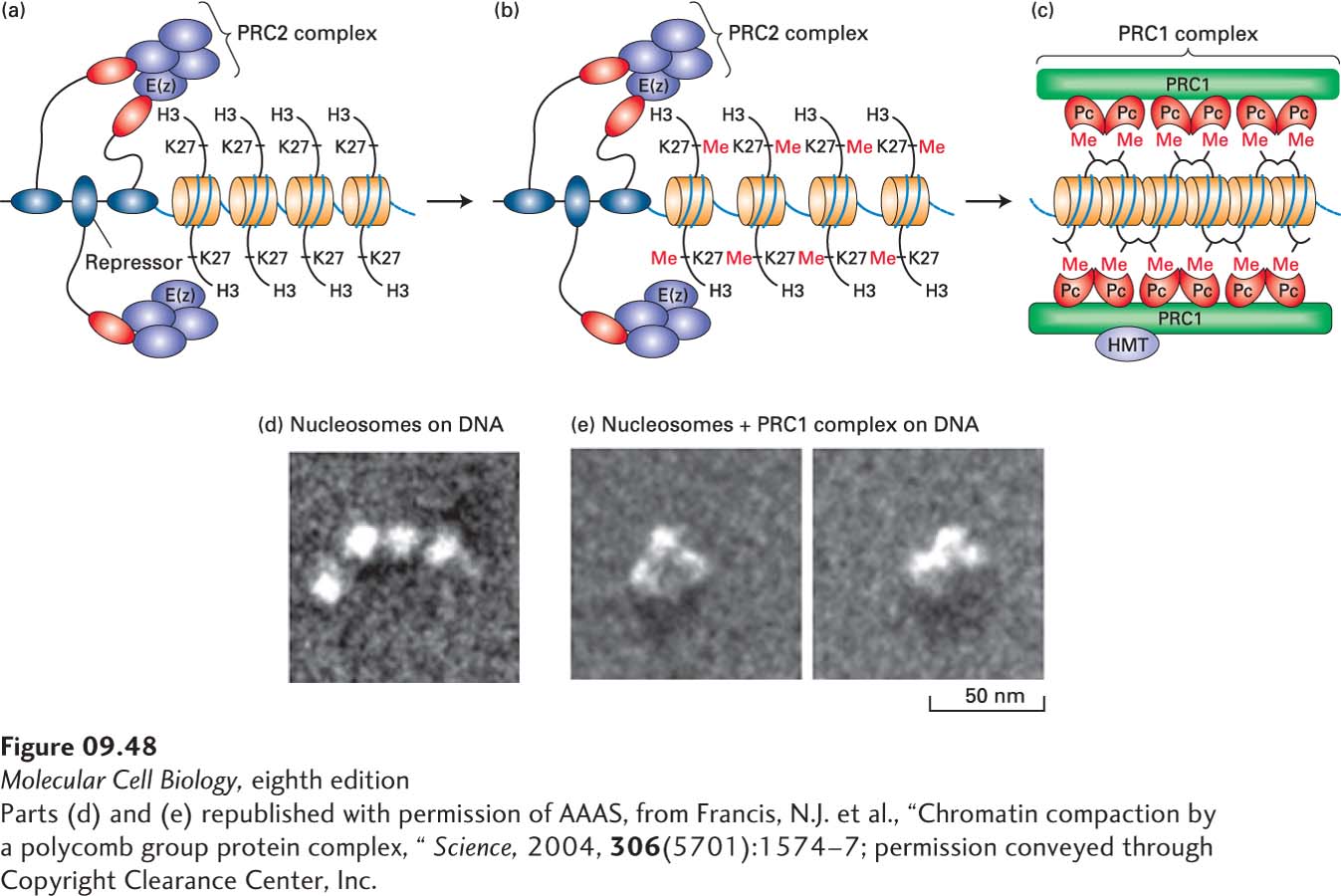
A current model for repression by Polycomb proteins is depicted in Figure 9-48. Most Polycomb proteins are subunits of one of two classes of multiprotein Polycomb repressive complexes: PRC1 and PRC2. The PRC2 complexes are thought to act initially by associating with the repression domains of specific repressors bound to their cognate DNA sequences early in embryogenesis, or with ribonucleoprotein complexes containing long noncoding RNAs, as discussed in a later section. The PRC2 complexes contain histone deacetylases that inhibit transcription, as discussed above. They also contain a subunit [E(z) in Drosophila, EZH2 in mammals] with a SET domain, which is the catalytic domain of several histone methyl transferases. This SET domain in PRC2 complexes methylates histone H3 on lysine 27, generating di- and trimethyl lysines. A PRC1 complex then binds the methylated nucleosomes through dimeric Pc subunits (CBXs in mammals), each containing a methyl lysine–binding domain (called a chromodomain) specific for methylated H3 lysine 27. Binding of the dimeric Pc to neighboring nucleosomes is proposed to condense the chromatin into a structure that inhibits transcription. This proposal is supported by electron microscopy studies showing that PRC1 complexes cause nucleosomes to associate in vitro (Figure 9-48d, e).
[Parts (d) and (e) republished with permission of AAAS, from Francis, N.J. et al., “Chromatin compaction by a polycomb group protein complex, “Science, 2004, 306(5701):1574–7; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 9-48Model for repression by Polycomb complexes. (a) During early embryogenesis, repressors associate with the PRC2 complex. (b) This association results in methylation (Me) of neighboring nucleosomes on histone H3 lysine 27 (K27) by the SET domain–containing subunit E(z). (c) The PRC1 complex binds nucleosomes methylated at H3 lysine 27 through a dimeric, chromodomain-containing subunit Pc. The PRC1 complex condenses the chromatin into a repressed chromatin structure. PRC2 complexes associate with PRC1 complexes to maintain H3 lysine 27 methylation of neighboring histones. As a consequence, PRC1 and PRC2 association with the region is maintained when expression of the repressor proteins in (a) ceases. (d, e) Electron micrograph of a 1-kb fragment of DNA bound by four nucleosomes in the absence (d) and presence (e) of one PRC1 complex per five nucleosomes. See A. H. Lund and M. van Lohuizen, 2004, Curr. Opin. Cell Biol.16:239; and N. J. Francis, R. E. Kingston, and C. L. Woodcock, 2004, Science306:1574.
[Parts (d) and (e) republished with permission of AAAS, from Francis, N.J. et al., “Chromatin compaction by a polycomb group protein complex, “Science, 2004, 306(5701):1574–7; permission conveyed through Copyright Clearance Center, Inc.]
PRC1 complexes also repress transcription through additional mechanisms. The PRC1 complex contains a ubiquitin ligase that monoubiquitinylates histone H2A at lysine 119 in the H2A C-terminal tail (see Figure 8-26). This modification of H2A inhibits elongation by inhibiting a histone chaperone that removes histone octamers from DNA as Pol II transcribes through a nucleosome, then replaces them as the polymerase passes. PRC1 also associates with a histone demethylase that specifically removes methyl groups from lysine 4 of histone H3, an activating mark discussed above.
PRC2 complexes associate with nucleosomes bearing the histone H3 lysine 27 trimethylation mark, maintaining methylation of H3 lysine 27 in nucleosomes in the region. This methylation results in association of the chromatin with PRC1 and PRC2 complexes even after expression of the initial repressor proteins shown in Figure 9-48a, b has ceased. This association maintains H3 lysine 27 methylation by a mechanism analogous to that diagrammed in Figure 9-47. This mechanism is a key feature of Polycomb repression, which is maintained through successive cell divisions for the life of an organism (~100 years for some vertebrates, 2000 years for a sugar cone pine!).
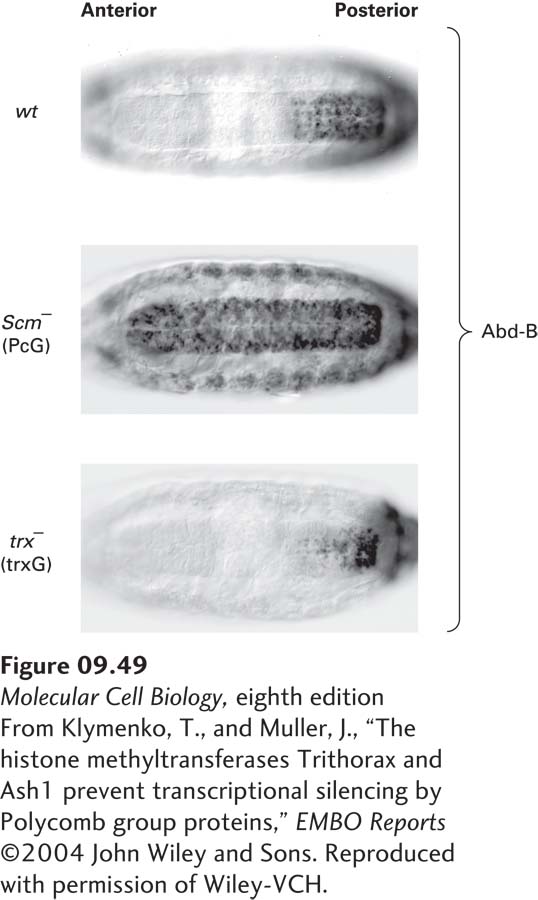
Trithorax proteins counteract the repressive mechanism of Polycomb proteins, as shown in studies of expression of the Hox transcription factor Abd-B in the Drosophila embryo (Figure 9-49). Abd-B is normally expressed only in posterior segments of the developing embryo. When the Polycomb system is defective, Abd-B is expressed in all cells of the embryo. When the Trithorax system is defective and cannot counteract repression by the Polycomb system, Abd-B is repressed in most cells, except those in the very posterior of the embryo. Trithorax complexes include a histone methyl transferase that trimethylates histone H3 lysine 4, a histone methylation that is associated with the promoters of actively transcribed genes. This histone modification creates a binding site for histone acetylase and for chromatin-remodeling complexes that promote transcription, as well as for TFIID, the general transcription factor that initiates preinitiation-complex assembly (see Figure 9-19). Nucleosomes with H3 lysine 4 methylation are also binding sites for specific histone demethylases that remove H3 histone K9 and K27 methylation, preventing the binding of HP1 and the Polycomb repressive complexes. Nucleosomes marked with H3 lysine 4 methylation are also thought to be distributed to both daughter DNA molecules during DNA replication, resulting in maintenance of this epigenetic mark by a strategy similar to that diagrammed in Figure 9-47.
FIGURE 9-49Opposing influence of Polycomb and Trithorax complexes on expression of the Hox transcription factor Abd-B in Drosophila embryos. At the stage of Drosophila embryogenesis shown, Abd-B is normally expressed only in posterior segments of the developing embryo, as shown at the top (wt) by immunostaining with a specific anti–Abd-B antibody. In embryos with homozygous mutations of Scm, a Polycomb gene (PcG) encoding a protein associated with the PRC1 complex, Abd-B expression is derepressed in all embryo segments. In contrast, in homozygous mutants of trx, a Trithorax gene (trxG), Abd-B repression is increased so that the protein is expressed at high concentrations only in the most posterior segment.