Clonal selection is the basis for antibody specificity.
A central question in immunology is how an individual can generate a great diversity of antibodies, each of which is specific for a given antigen. There are seemingly limitless types of antigen that an organism might come across. How can B cells generate antibodies that interact only with specific antigens among this tremendous diversity?
Two alternative hypotheses were suggested to explain antibody specificity. In 1900, Paul Ehrlich suggested that there exists a large pool of B cells, each with a different antibody on its surface. The antigen binds to one or a few of these antibodies, stimulating the B cell to divide. In this model, specificity exists before any exposure to the antigen. Ehrlich’s model drew on concepts from chemistry such as the binding of an enzyme to a substrate in the manner of a lock and key (Chapter 6).
Later, in the early 1900s, a different hypothesis was suggested. This hypothesis proposed that the antigen instructs the antibody to fold in a particular way so that the two interact in a specific manner. Here, the antigen plays a more active role in the process, and antibody specificity is determined only after interaction with the antibody.
Which of these two hypotheses is correct? Does the antigen select a B cell with a preexisting surface antibody with which it interacts specifically, as in the first hypothesis, or does it instruct the B cell to make a specific antibody, as in the second?
Experimental data on the genetic basis for antibody diversity (described below) provided strong evidence for the first hypothesis. Every individual has a very large pool of B cells, each with a different antibody on its cell surface (Fig. 43.11). Collectively, these antibodies recognize a great diversity of antigens, but each individual antibody can recognize only one antigen or a few antigens. An antigen interacts with the antibody on the B cell, or a small set of B cells, that has the best fit for the antigen. Binding leads the B cell to divide and differentiate into two types of cell. Plasma cells are daughters of the B cell that secrete antibodies that bind to the antigen and elicit an immune response against the pathogen. Memory cells are long-
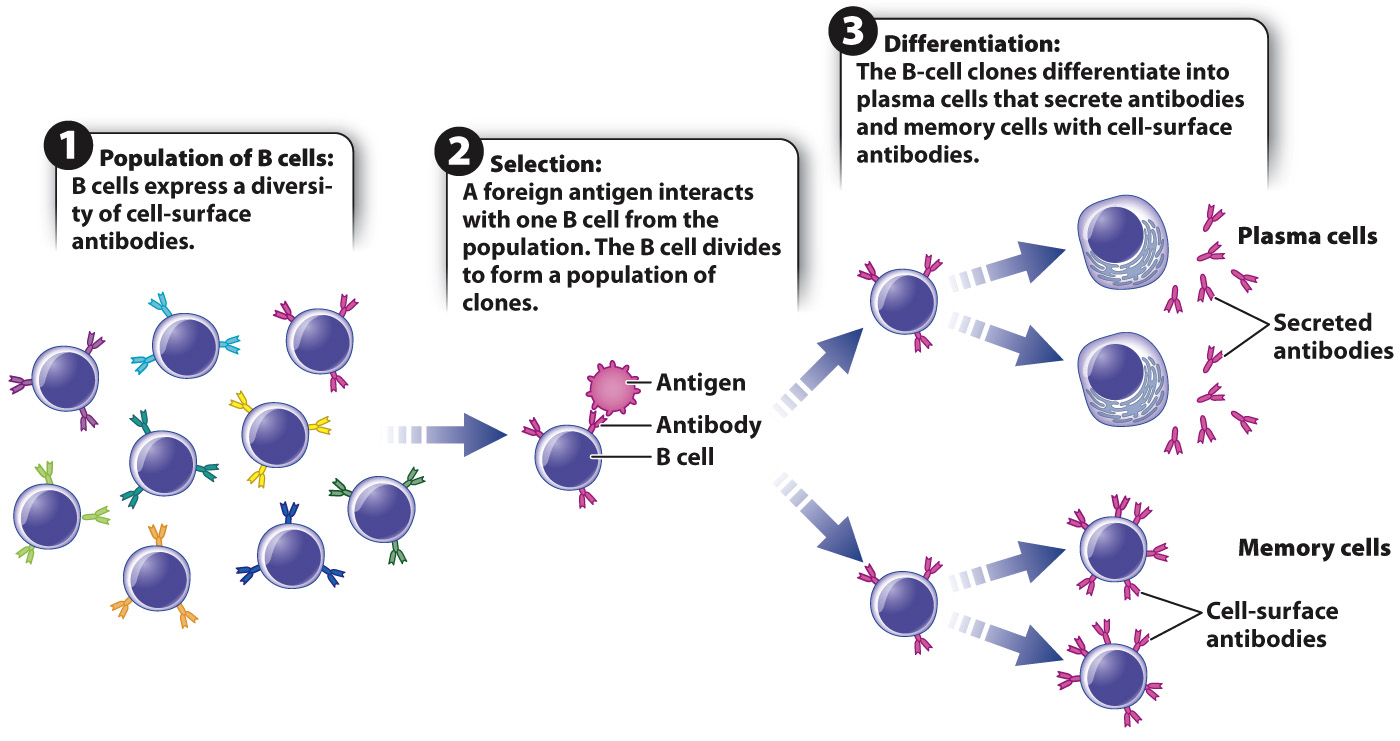
The process by which antigen binding generates a clone of B cells is called clonal selection, and it is one of the central principles of immunology. Almost counterintuitively, antibody specificity is achieved before any exposure to the antigen. In this way, clonal selection resembles natural selection (Chapter 21). In natural selection, there is genetic variation in a population and the environment selects among this variation, allowing some individuals to survive and reproduce more than others. Similarly, in clonal selection, there is variation in a population of B cells, and the antigen selects among this variation, allowing some cells to proliferate more than others.