The signals that regulate cell division fall into two basic types: molecules that stimulate cell division and molecules that inhibit it. These control mechanisms are similar to the accelerator and brake of a car. In normal cells (but, one would hope, not in your car), both accelerators and brakes are applied at the same time, causing cell division to proceed at the proper speed.
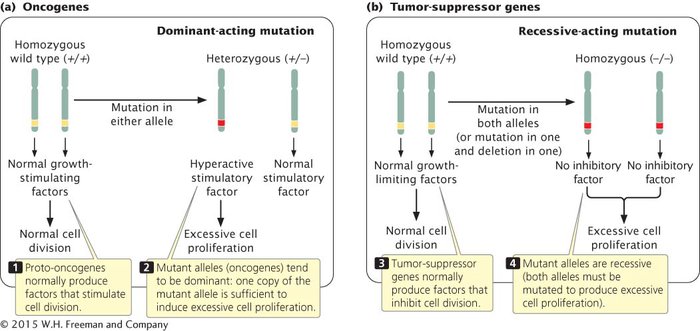
Because cell division is affected by both accelerators and brakes, cancer can arise from mutations in either type of signal, and thus there are several fundamentally different routes to cancer. A stimulatory gene can be made hyperactive, or active at inappropriate times, which is analogous to having a car’s accelerator stuck in the floored position. Mutations in stimulatory genes usually act in a dominant manner because even the amount of gene product produced by a single allele is usually sufficient to have a stimulatory effect. Mutated copies of dominant-acting stimulatory genes that cause cancer are termed oncogenes (Figure 16.5a).
16.5 Both oncogenes (a) and tumor-suppressor genes (b) contribute to cancer, but differ in their modes of action and dominance.
Cell division may also be stimulated when inhibitory genes are made inactive, which is analogous to having a defective brake in a car. Mutated inhibitory genes generally act in a recessive manner because both copies must be mutated to remove all inhibition. Inhibitory genes involved in cancer are termed tumor-suppressor genes (Figure 16.5b). Many cancer cells have mutations in both oncogenes and tumor-suppressor genes.
Although oncogenes or mutated tumor-suppressor genes, or both, are required to produce cancer, mutations in DNA-repair genes can increase the likelihood of acquiring mutations in those genes. Having mutated DNA-repair genes is analogous to having a lousy car mechanic who does not make the necessary repairs on a broken accelerator or brake.
ONCOGENES Oncogenes were the first type of cancer-causing genes to be identified. In 1909, a farmer brought physician Peyton Rous a hen with a large connective-tissue tumor (sarcoma) growing on its breast. When Rous injected pieces of this tumor into other hens, they also developed sarcomas. Rous conducted experiments that demonstrated that the tumors were being transmitted by a retrovirus, which became known as the Rous sarcoma virus, as mentioned in Chapter 7. A number of other cancer-causing viruses were subsequently isolated from various animal tissues. These viruses were generally assumed to carry a cancer-causing gene that was transferred to the host cell. The first oncogene, called src, was isolated from the Rous sarcoma virus in 1970.
In 1975, Michael Bishop, Harold Varmus, and their colleagues began to use probes for viral oncogenes to search for related sequences in normal cells. They discovered that the genomes of all normal cells carry DNA sequences that are closely related to oncogenes. These normal cellular genes are called proto-oncogenes. They are responsible for basic cellular functions in normal cells, but when mutated, they become oncogenes that contribute to the development of cancer. When a virus infects a cell, a proto-oncogene may become incorporated into the viral genome through recombination. Within the viral genome, the proto-oncogene may mutate to an oncogene that, when inserted into a host cell, causes rapid cell division and cancer. Because the proto-oncogenes are more likely to undergo mutation or recombination within a virus than within the host cell, viral infection is often associated with some cancers.
Page 433
Proto-oncogenes can be converted into oncogenes in viruses in several different ways. The sequence of the proto-oncogene may be altered or truncated as it is incorporated into the viral genome. This mutated copy of the gene may then produce an altered protein in the host cell that causes uncontrolled cell proliferation. Alternatively, through recombination, a proto-oncogene may end up next to a viral promoter or enhancer, which causes the gene to be overexpressed in the host cell. Finally, the function of a proto-oncogene in the host cell may be altered when a virus inserts its own DNA into the gene, disrupting its normal function. While viruses are capable of converting proto-oncogenes into oncogenes, most proto-oncogenes are mutated to form oncogenes without the involvement of a virus.
Many oncogenes have been identified by experiments in which selected fragments of DNA are added to cells in culture. Some of the cells take up the DNA, and if these cells become cancerous, then the DNA fragment that was added to the culture must contain an oncogene. The fragments can then be sequenced and the oncogene identified. A large number of oncogenes have now been discovered (Table 16.4). About 90% of all cancer genes are thought to be dominant oncogenes.
TABLE 16.4Some oncogenes and functions of their corresponding proto-oncogenes
Gene
Normal function
Cancer in which gene is mutated
erbB
Part of growth factor receptor
Many types of cancer
fos
Transcription factor
Osteosarcoma and endometrial carcinoma
jun
Transcription factor, cell cycle control
Lung cancer, breast cancer
myc
Transcription factor
Lymphomas, leukemias, neuroblastoma
ras
GTP binding and GTPase
Many types of cancer
sis
Growth factor
Glioblastomas and other cancers
src
Protein tyrosine kinase
Many types of cancer
TUMOR-SUPPRESSOR GENES Tumor-suppressor genes are more difficult to identify than oncogenes because they inhibit cancer and are recessive; both alleles must be mutated before the inhibition of cell division is removed. Because it is the failure of their function that promotes cell proliferation, tumor-suppressor genes cannot be identified by adding them to cells and looking for cancer. About 10% of cancer-causing genes are thought to be tumor-suppressor genes.
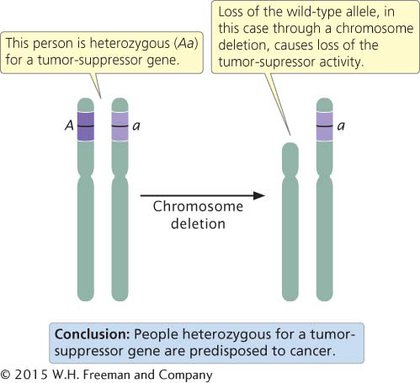
Defects in both copies of a tumor-suppressor gene are usually required to cause cancer. An organism can inherit one defective copy of the tumor-suppressor gene (be heterozygous for the cancer-causing mutation) and not have cancer because the remaining normal allele produces the tumor-suppressing product. However, these heterozygotes are often predisposed to cancer because the inactivation or loss of that one remaining allele is all that is required to completely eliminate the tumor-suppressor product. Inactivation of the remaining wild-type allele in heterozygotes is referred to as the loss of heterozygosity. A common mechanism for the loss of heterozygosity is a deletion on the chromosome that carried the normal copy of the tumor-suppressor gene (Figure 16.6).
Page 434
16.6 The loss of heterozygosity often leads to cancer in a person heterozygous for a tumor-suppressor gene.
Among the first tumor-suppressor genes to be identified was the gene that causes retinoblastoma. In 1985, Raymond White and Webster Cavenne showed that large segments of chromosome 13 were missing in cells from retinoblastoma tumors, and later, the tumor-suppressor gene was isolated from those segments. Another example of a tumor-suppressor gene is BRCA1, mutations of which are associated with increased risk of breast and ovarian cancer. BRCA1 produces a protein that normally helps in the repair of double-strand breaks in DNA by homologous recombination. That protein also acts as a transcription factor and interacts with histone deacetylase enzymes, which affect transcription. A number of tumor-suppressor genes have now been discovered (Table 16.5).
TABLE 16.5Some tumor-suppressor genes and their normal functions
Gene
Normal function
Cancer in which gene is mutated
APC
Scaffold protein, interacts with microtubules
Colorectal
CDKN2A
Regulates cell division
Melanoma
BRCA1
DNA repair, transcription factor
Breast and ovarian
NF1
GTPase activator
Neurofibromatosis
p53
Regulates cell division
Many types of cancer
RB
Regulates cell division
Retinoblastoma
Sometimes the mutation or loss of a single allele of a recessive tumor-suppressor gene is sufficient to cause cancer. This effect—the appearance of the trait in an individual cell or organism that is heterozygous for a normally recessive trait—is called haploinsufficiency. This phenomenon is thought to be due to dosage effects: the heterozygote produces only half as much of the product encoded by the tumor-suppressing gene as the homozygote does. Normally, this amount is sufficient for the cellular processes that prevent tumor formation, but it is less than the optimal amount, and other factors may sometimes combine with the lowered amount of tumor-suppressor product to cause cancer. TRY PROBLEM 19
CONCEPTS
Proto-oncogenes are genes that control normal cellular functions; when mutated, they become oncogenes that stimulate cell proliferation. They tend to be dominant in their action. Tumor-suppressor genes normally inhibit cell proliferation; when mutated, they allow cells to proliferate. Tumor-suppressor genes tend to be recessive in their action. Individual organisms that are heterozygous for tumor-suppressor genes are often predisposed to cancer.
CONCEPT CHECK 2
Why are oncogenes usually dominant in their action, whereas tumor-suppressor genes are recessive?
Oncogenes stimulate cell proliferation. Mutations in oncogenes are usually dominant because a mutation in a single copy of the gene is usually sufficient to produce a stimulatory effect. Tumor-suppressor genes inhibit cell proliferation. Mutations in tumor-suppressor genes are generally recessive because both copies must be mutated to remove all inhibition.
 TRY PROBLEM 19
TRY PROBLEM 19 CONCEPT CHECK 2
CONCEPT CHECK 2