CONCEPT9.1 DNA Structure Reflects Its Role as the Genetic Material
In Chapter 8 we described Mendel’s experiments in the 1860s demonstrating that genes are physically distinct entities, and other work in the early twentieth century showing that groups of genes are linked together. By the early twentieth century, a “chromosomal theory of inheritance” had been developed, proposing that Mendel’s genes are present in the chromosomes of the cell nucleus. This theory came partly from observations of sea urchins: it was shown that an entire set of chromosomes must be present for a sea urchin embryo to grow and develop. Scientists also observed that homologous chromosomes are paired during meiosis, that crossing over occurs during meiosis I (see Figure 8.14), and that the chromosome pairs separate independently at anaphase I. Thus the behavior of chromosomes accounted for Mendel’s laws of segregation and independent assortment, as well as the later discoveries of linkage and recombination.
We now turn to the actual chemical nature of genes, beginning with the evidence that DNA is the carrier of heritable information. Scientists used two types of evidence to show that DNA is the genetic material: circumstantial and experimental. We will provide examples of both types.
Circumstantial evidence suggested that DNA is the genetic material
Early observations pointed to the possibility that DNA is the genetic material. Scientists found that DNA:
- is present in the cell nucleus and in condensed chromosomes
- doubles during S phase of the cell cycle
- is twice as abundant in the diploid cells as in the haploid cells of a given organism.
Let’s look at some of these lines of evidence.
DNA in The Nucleus
DNA was first isolated in 1868 by the young Swiss researcher–physician Friedrich Miescher, who isolated cell nuclei from white blood cells in pus from the bandages of wounded soldiers. When he treated these nuclei chemically, a fibrous substance came out of solution. He called it “nuclein” and found that it contained the elements C, H, O, N, and P. With no evidence except for finding it in the nucleus, Miescher boldly proposed that nuclein was the genetic material. His supervising professor was so astounded by Miescher’s work that he repeated it himself in the laboratory, and finally allowed his student to publish it in a scientific journal.
DNA in The Chromosomes
In the early twentieth century dyes were developed that react specifically with DNA, only showing color when they bind to it. This allowed individual cells to be examined for the location and amount of DNA they contained. When dividing cells were stained with such a dye, only the chromosomes were stained:
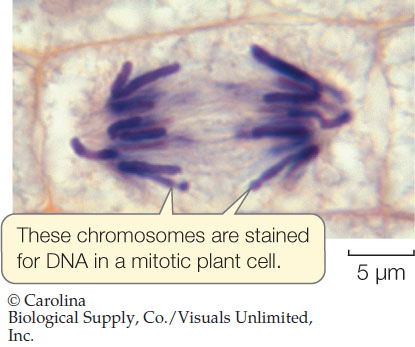
DNA Amounts
The amount of dye binding to DNA, and hence the intensity of color observed, was directly related to the amount of DNA present: the greater the intensity, the more DNA. This allowed scientists to analyze DNA amounts in individual cells during the cell cycle (see Concept 7.2).
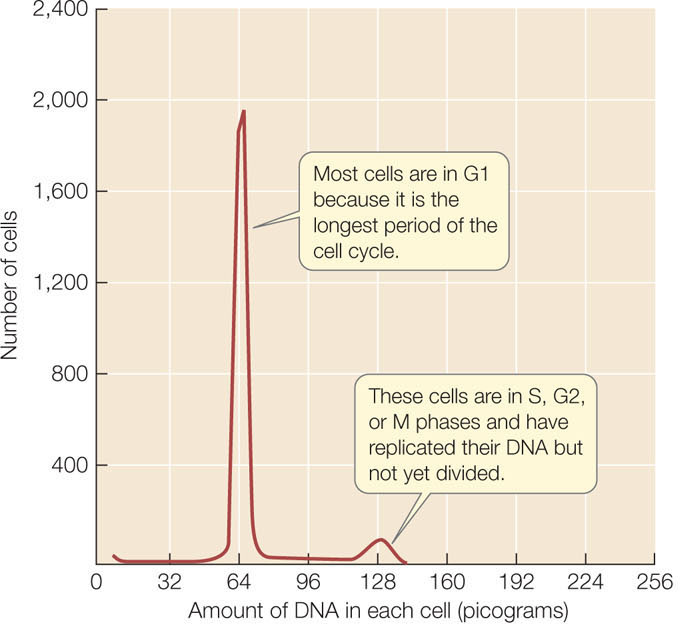
When a population of actively dividing cells was stained with dye, the amount of DNA in each cell could be quantified by passing the cells one by one through an instrument called a flow cytometer. In general, two populations of cells were seen: most cells were in G1 and contained half the amount of DNA that was in the remaining cells, which were in S, G2, or M (FIGURE 9.1). Such staining experiments confirmed two other predictions for DNA as the genetic material:
- Virtually all nondividing somatic cells of a particular organism have the same amount of nuclear DNA. This amount varies from species to species.
- Similar experiments showed that after meiosis, gametes have half the amount of nuclear DNA as somatic cells.
174
Experimental evidence confirmed that DNA is the genetic material
Circumstantial evidence can show correlations between two phenomena. However, scientists rely on experiments to provide evidence of a cause-and-effect relationship. Chromosomes in eukaryotic cells contain DNA, but they also contain proteins that are bound to DNA. Therefore it was difficult to rule out the possibility that genetic information might be carried in proteins. In order to confirm that DNA was the genetic material, biologists used model organisms such as bacteria in transformation experiments. They found, for example, that the addition of DNA from one strain of bacterium could genetically transform another strain:
Bacterium strain A + strain B DNA → bacterium strain B
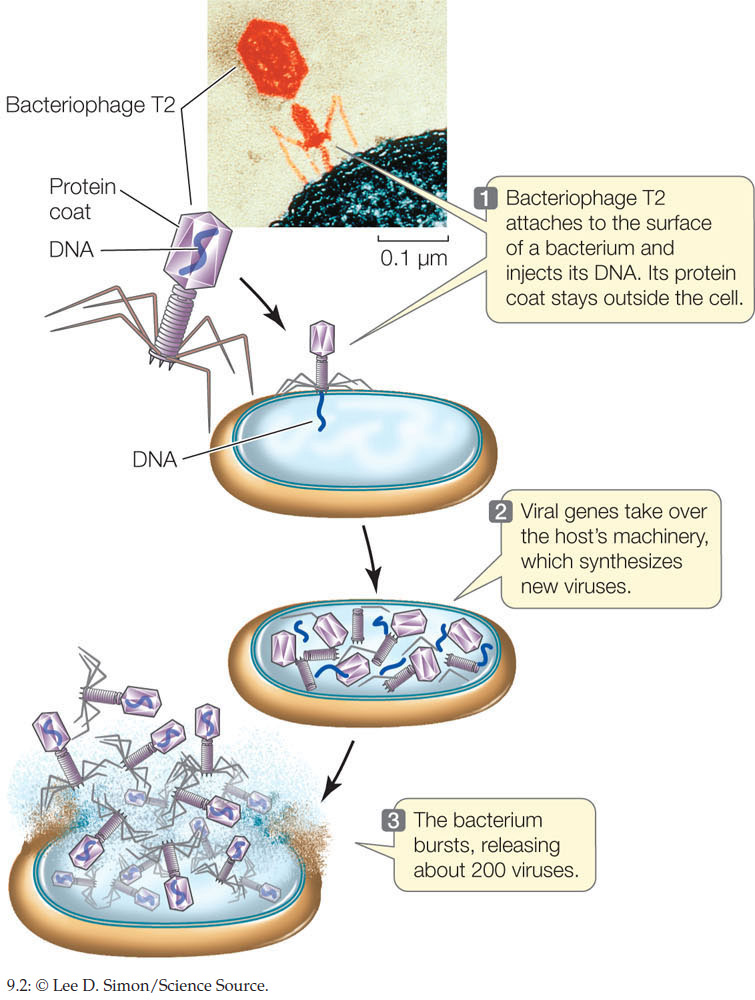
Viruses provided another system to explore this question. Many viruses, including bacteriophage (viruses that infect bacteria), are composed of DNA and only one or a few kinds of protein. When a bacteriophage infects a bacterium, it takes about 20 minutes for the virus to hijack the bacterium’s metabolic capabilities and turn the bacterium into a virus factory. Minutes later, the bacterium is dead and hundreds of viruses are released.
The transition from bacterium to virus producer is a change in the genetic program of the bacterial cell, resulting in a change of phenotype. Experiments showed that only the viral DNA is injected into the cell during infection (FIGURE 9.2). Since the viral DNA genetically transformed the bacteria, this was further evidence that DNA and not protein is the genetic material.
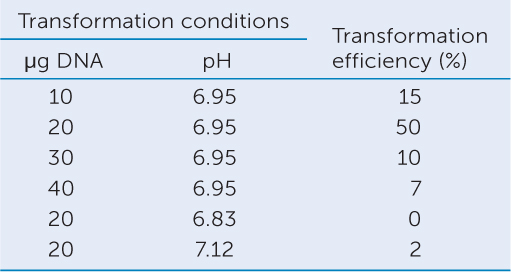
The transformation of mammalian cells with a gene for antibiotic resistance provided another model system for showing that DNA is the genetic material (FIGURE 9.3). When cultured mammalian cells were treated with DNA containing a gene for resistance to the antibiotic neomycin, the cells were able to grow on media containing the antibiotic.
Investigation
HYPOTHESIS
DNA can transform eukaryotic cells.
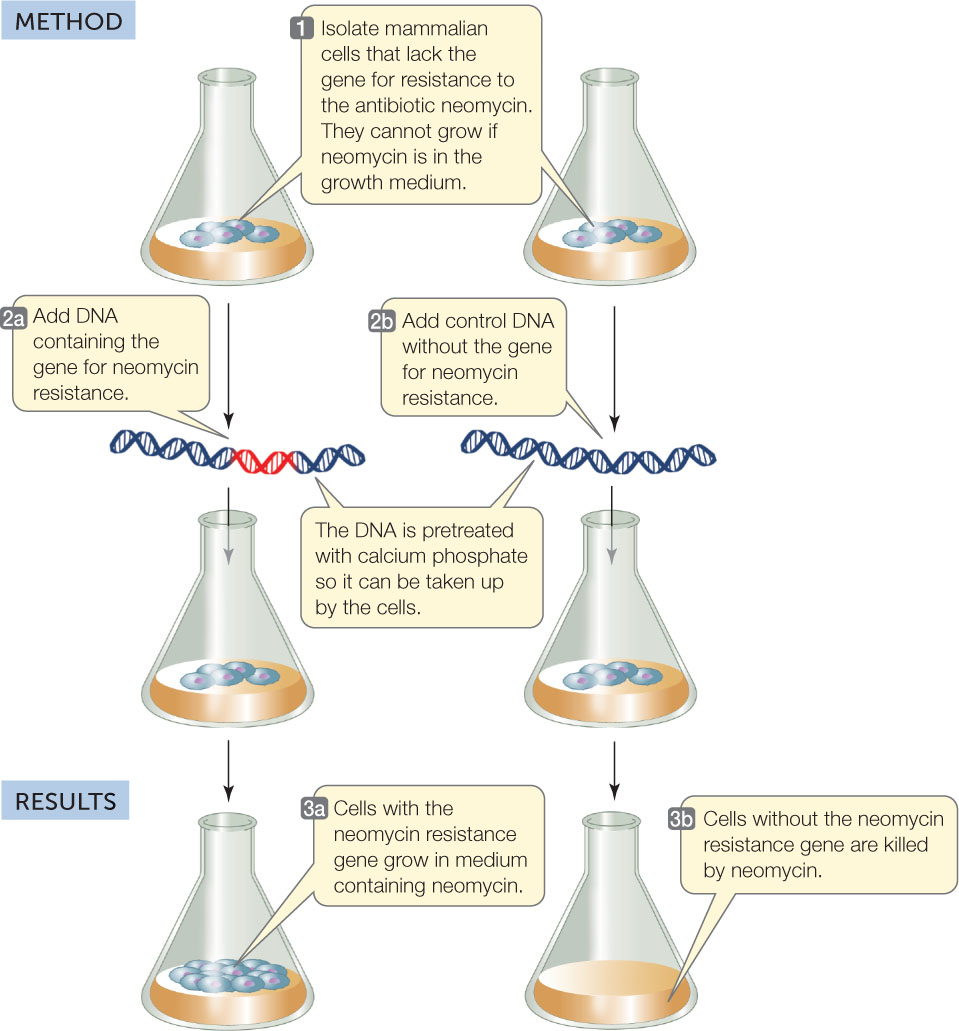
CONCLUSION
The cells were transformed by DNA.
ANALYZE THE DATA
Transformation was achieved by adding the DNA in a solution of calcium phosphate (Ca3[PO4]2) at pH 6.95. Ca3(PO4)2 produces Ca2+ in solution; this neutralizes negative charges on the DNA and on the cell membrane, thus allowing the DNA to pass through the membrane. In other experiments, the type or amount of DNA and pH were varied. Transformation efficiency was calculated as the percentage of cells that produced colonies on a medium containing neomycin, compared with cells growing on medium without neomycin. Explain the transformation efficiency in terms of the conditions given in the data.
a C. Chen and H. Okayama. 1987. Molecular and Cellular Biology 7: 2745–2752.
Many kinds of cells can be transformed in this way—even egg cells. In this case, a whole new genetically transformed organism can result. The fertilized egg can develop into a new multicellular organism through mitosis; such an organism is referred to as transgenic. These methods form the basis of much applied research, including biotechnology and genetic engineering. The transformation of multicellular eukaryotes provides powerful experimental evidence for DNA as the genetic material.
The Discovery Of The Three-Dimensional Structure Of DNA Was A Milestone In Biology
Mendel showed that genes are physically distinct entities, and further research identified DNA as the genetic material. The history of how the actual structure of DNA was deciphered is worth considering, as it represents not only talented scientists working together, but also a landmark in our understanding of biology.
By the mid-twentieth century, the chemical makeup of DNA, as a polymer made up of nucleotide monomers, had been known for several decades. In determining the structure of DNA, scientists hoped to answer two additional questions:
- How is DNA replicated between cell divisions?
- How does it direct the synthesis of specific proteins?
They were eventually able to answer both questions. The structure of DNA was deciphered only after many types of experimental evidence were considered together.
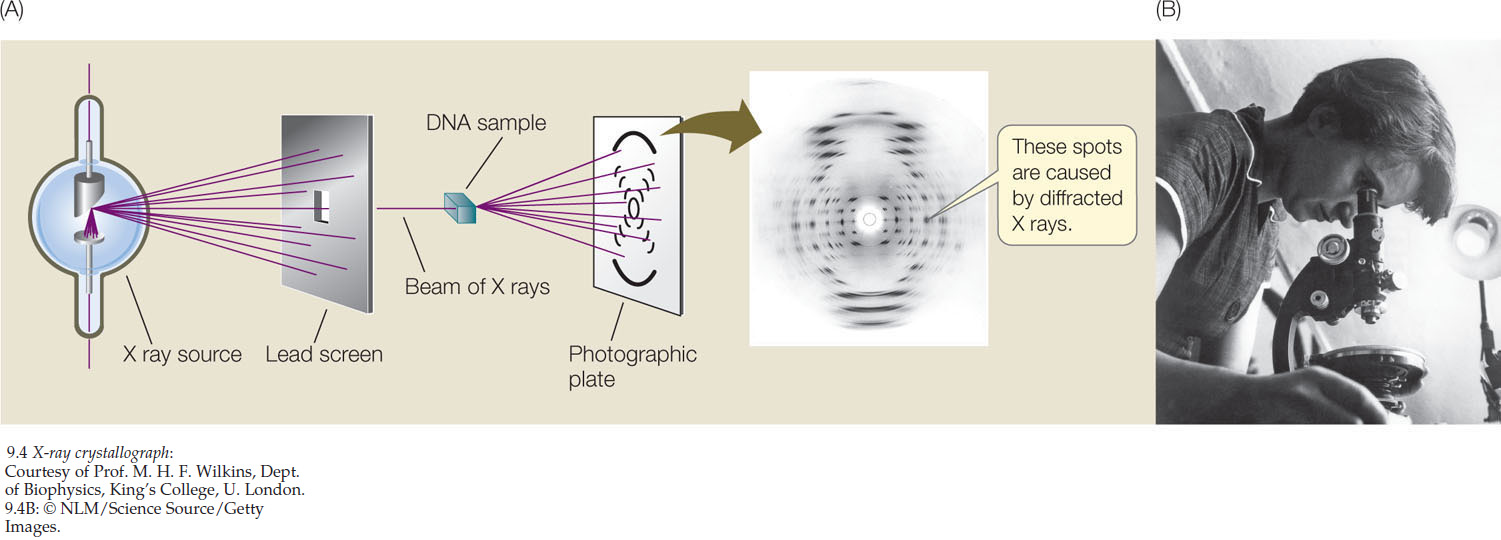
X-Ray Crystallography Provided Clues To DNA’s Structure
The most crucial evidence was obtained using X-ray crystallography. Some chemical substances, when they are isolated and purified, can be made to form crystals. The positions of atoms in a crystallized substance can be inferred from the diffraction pattern of X rays passing through the substance (FIGURE 9.4A). The structure of DNA would not have been characterized without the crystallographs prepared in the early 1950s by the English chemist Rosalind Franklin (FIGURE 9.4B). Franklin’s work, in turn, depended on the success of the English biophysicist Maurice Wilkins, who prepared samples containing very uniformly oriented DNA fibers. These fibers and the crystallographs Franklin prepared from them suggested a spiral or helical molecule.
The Nucleotide Composition Of DNA Was Known
The chemical composition of DNA also provided important clues to its structure. Biochemists knew that DNA is a polymer of nucleotides. Each of these nucleotides consists of a molecule of the sugar deoxyribose, a phosphate group, and a nitrogen-containing base (see Figure 3.1). The only differences among the four nucleotides of DNA are their nitrogenous bases: the purines adenine (A) and guanine (G), and the pyrimidines cytosine (C) and thymine (T) (see Figure 3.1).
175
176
In 1950, biochemist Erwin Chargaff at Columbia University reported an important observation. He and his colleagues had found that DNA samples from many different species—and from different sources within a single organism—exhibited certain regularities. The following rule held for each sample: the amount of adenine equaled the amount of thymine (A = T), and the amount of guanine equaled the amount of cytosine (G = C). As a result, the total abundance of purines (A + G) equaled the total abundance of pyrimidines (T + C):
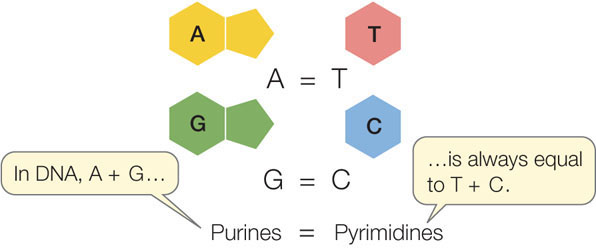
The structure of DNA could not have been worked out without this observation, now known as Chargaff’s rule.
Watson And Crick Described The Double Helix
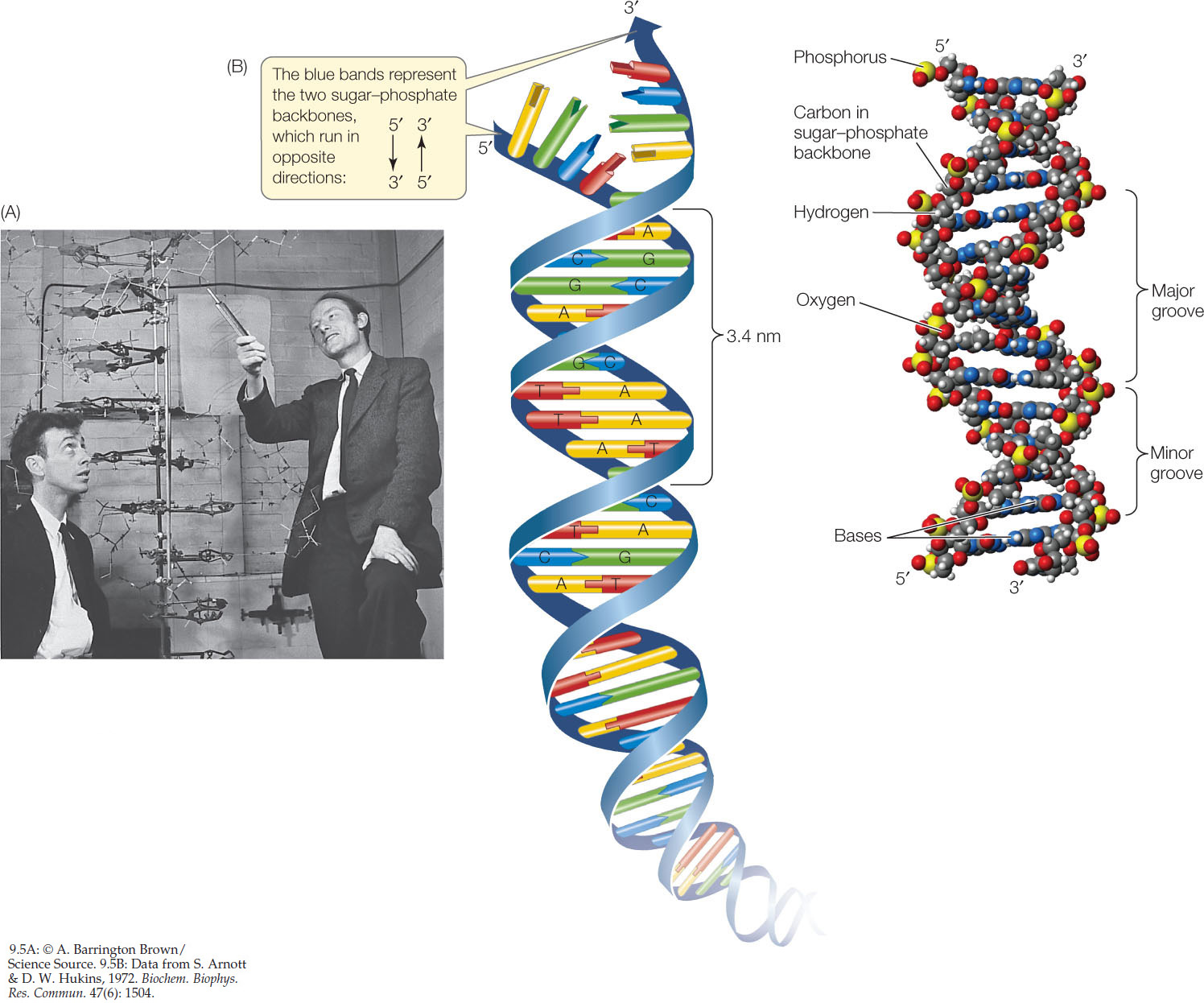
Chemical model building is the assembly of three-dimensional structures using known relative molecular dimensions and known bond angles. The English physicist Francis Crick and the American geneticist James D. Watson (FIGURE 9.5A), both then at the Cavendish Laboratory of Cambridge University, used model building to solve the structure of DNA. Rosalind Franklin’s crystallography results convinced them that the DNA molecule must be helical—it must have a spiral shape like a spring. Density measurements and previous model building experiments suggested that there are two polynucleotide chains in the molecule. Modeling studies also showed that the strands run in opposite directions, that is, they are antiparallel. The two strands would not fit together in the model if they were parallel.
How are nucleotides oriented in DNA chains? Watson and Crick suggested that:
- the nucleotide bases are on the interior of the two strands, with a sugar-phosphate backbone on the outside. The strands would not fit together otherwise:
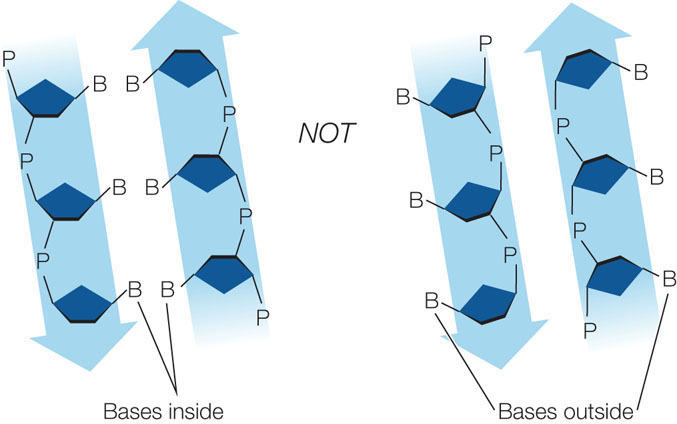
- to satisfy Chargaff’s rule (purines = pyrimidines), a purine on one strand is always paired with a pyrimidine on the opposite strand. These base pairs (A-T and G-C) have the same width down the double helix, a uniformity shown by X-ray diffraction:
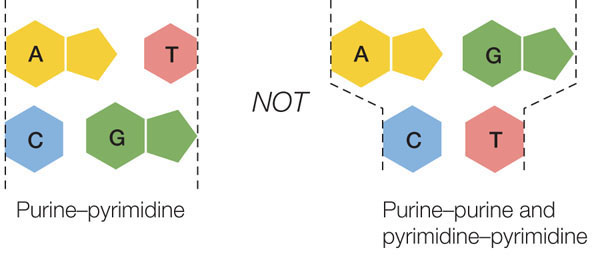
In late February of 1953, Crick and Watson built a model out of tin that established the general structure of DNA. This structure explained all the known chemical properties of DNA, and it opened the door to understanding its biological functions. There have been minor amendments to that first published structure, but its principal features remain unchanged.
Four key features define DNA structure
Four features summarize the molecular architecture of the DNA molecule (FIGURE 9.5B; also review Figure 3.4):
- DNA is a double-stranded helix of uniform diameter. The sugar–phosphate backbones of the two chains form a coil on the outside of the helix, and the nitrogenous bases point toward the center. The chains are held together by two chemical forces: hydrogen bonding between the basesand van der Waals forces between adjacent bases on the same strand. When the base rings come near one another, they tend to stack like poker chips because of these weak attractions.

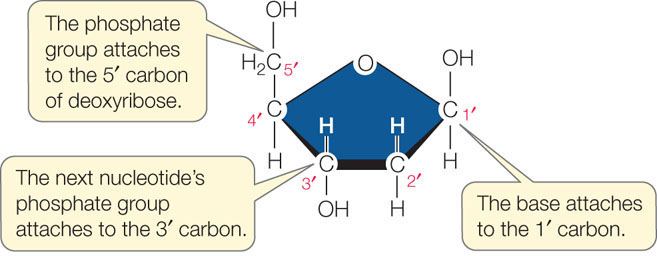
- The two DNA strands are antiparallel. The backbone of each strand contains repeating units of the monosaccharide (sugar) deoxyribose:The number followed by the prime sign (′) designates the position of a carbon atom in the sugar. In the sugar–phosphate backbone of DNA, the phosphate groups are connected to the 5′ carbon of one deoxyribose molecule and to the 3′ carbon of the next, linking the two sugar molecules together. Thus the two ends of the polynucleotide chain are different. The 5′ end of the chain is a free (not connected to another nucleotide) phosphate group. The 3′ end of the chain is a free 3′ hydroxyl (OH) group. In the double helix of DNA, the 5′ end of one strand is paired with the 3′ end of the other strand, and vice versa (see Figure 3.4).
- In DNA, the outer edges of the nitrogenous bases are exposed in the major and minor grooves. These grooves exist because the helices formed by the backbones of the two DNA strands are not evenly spaced relative to one another (see Figure 9.5B). FIGURE 9.6 shows the four possible configurations of the flat, hydrogen-bonded base pairs within the major and minor grooves. The exposed outer edges of the base pairs are accessible for additional hydrogen bonding. Notice that the arrangements of unpaired atoms and groups differ in the A-T base pairs compared with the G-C base pairs. Thus the surfaces of the A-T and G-C base pairs are chemically distinct, allowing other molecules, such as proteins, to recognize specific base-pair sequences and bind to them. The atoms and groups in the major groove are more accessible, and tend to bind other molecules more frequently, than those in the minor groove. This binding of proteins to specific base-pair sequences is the key to protein—DNA interactions, which are necessary for the replication and expression of the genetic information in DNA.
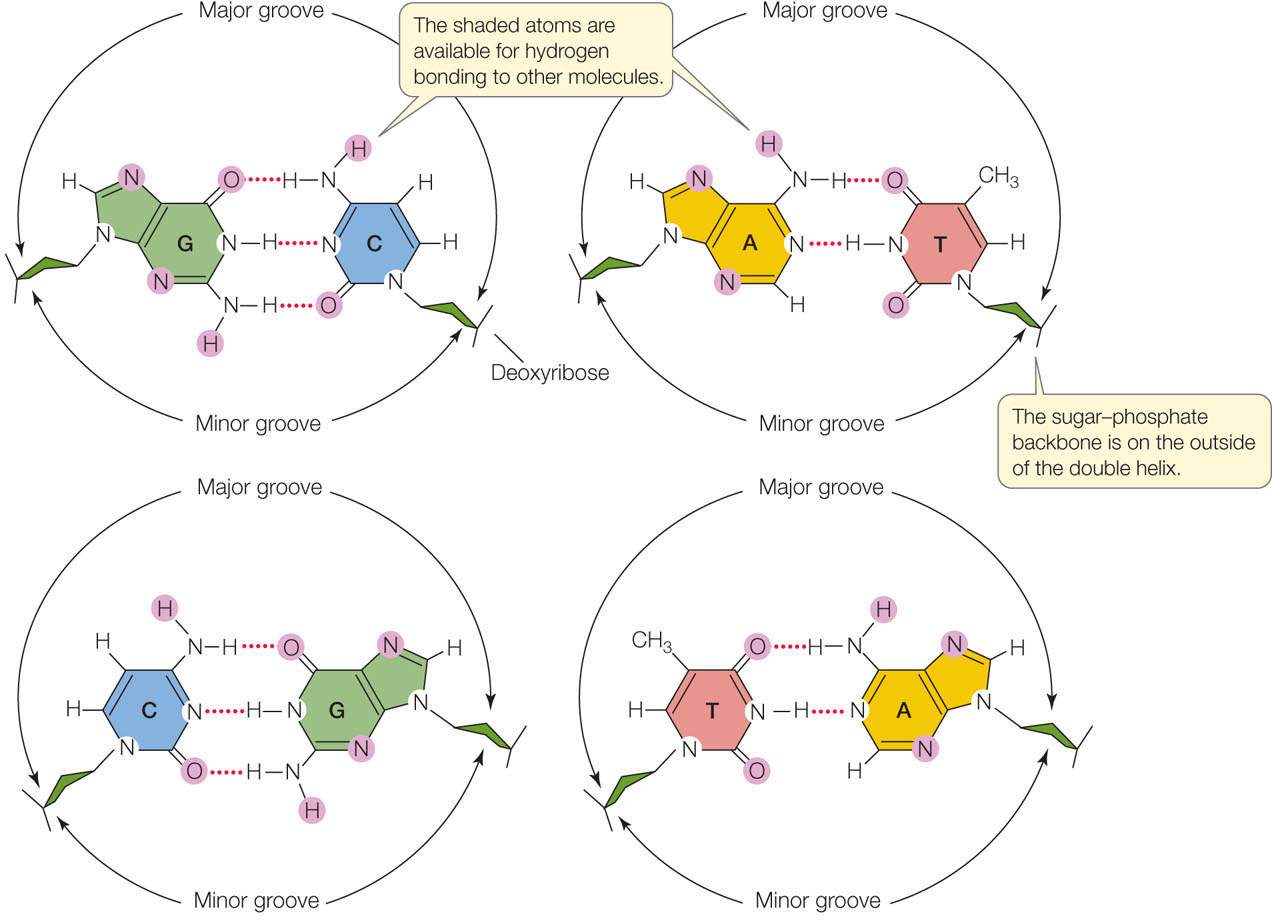 Figure 9.6: Base Pairs in DNA Can Interact with Other Molecules These diagrams show the four possible configurations of base pairs within the double helix. Atoms shaded in purple are available for hydrogen bonding with other molecules. These differences in configuration allow proteins to recognize specific DNA sequences.
Figure 9.6: Base Pairs in DNA Can Interact with Other Molecules These diagrams show the four possible configurations of base pairs within the double helix. Atoms shaded in purple are available for hydrogen bonding with other molecules. These differences in configuration allow proteins to recognize specific DNA sequences. - The DNA double helix is right-handed. Hold your right hand with the thumb pointing up (see Figure 9.10B). Imagine the curve of the helix following the direction of your fingers as it winds upward, and you have the idea.
While DNA usually forms a right-handed helix, it can sometimes be found as a much less stable left-handed helix. So-called Z-DNA (“zig-zag DNA”) does not have major and minor grooves and is more elongated and less compact than normal DNA. Z-DNA appears to form in regions of DNA that are being actively transcribed, and it may play a role in stabilizing the DNA during transcription.
The double-helical structure of DNA is essential to its function
The genetic material performs four important functions, and the DNA structure proposed by Watson and Crick was elegantly suited to three of them.
178
- Storage of genetic information. With its millions of nucleotides, the base sequence of a DNA molecule can encode and store an enormous amount of information. Variations in DNA sequences can account for differences among species and individuals.
- Precise replication during the cell division cycle. Replication could be accomplished by complementary base pairing, A with T and G with C. In the original publication of their findings in 1953, Watson and Crick coyly pointed out, “It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.” We will turn to DNA replication in Concept 9.2.
- Susceptibility to mutations. The structure of DNA suggested an obvious mechanism for mutations—stable changes in the genetic material. Mutations might be simple changes in the linear sequence of base pairs. We will discuss mutations in Concept 9.3.
- Expression of the coded information as phenotypes. The way this function is accomplished is not obvious in the structure of DNA. However, as we described briefly in Chapter 3 (see Figure 3.5), the nucleotide sequence of DNA can be copied—transcribed—into RNA. The linear sequence of nucleotides that is RNA is then translated into a linear sequence of amino acids that can fold into a protein (see Figure 3.7). Thus the information carried in DNA is ultimately expressed as proteins. The many and varied enzymes and structural proteins encoded by an organism’s DNA determine most of the phenotypes of that organism.
LINK
The transcription of DNA into RNA and the translation of RNA into protein are described in detail in Concepts 10.2 and 10.4
CHECKpointCONCEPT9.1
- Compare circumstantial evidence with experimental evidence in science, using DNA as an example.
- If a DNA molecule is 20 percent G, what are the percentages of the other three bases?
- A single-stranded sequence of DNA contains 20 A’s, 25 G’s, 35 C’s, and 24 T’s. What is the total base composition of the double-stranded molecule?
- What circumstantial and experimental evidence would show that protein and not DNA is the genetic material? How would proteins differ from nucleic acids as informational molecules?
We have seen that a DNA molecule consists of long polymers of nucleotides. An individual DNA strand contains thousands or millions (up to about 1 billion) nucleotides in a precise sequence. How is this huge amount of genetic information replicated before cell division?
179