CONCEPT24.1 The Plant Body Is Organized and Constructed in a Distinctive Way
Plants must harvest energy from sunlight and mineral nutrients from the soil. Because they are stationary (sessile), they cannot move to find a more favorable environment or to avoid predators. Their body plan and physiology allow plants to respond to these challenges:
- Stems, leaves, and roots have structural adaptations that enable plants anchored to one spot to capture scarce resources effectively, both above and below the ground.
- Their ability to grow throughout their lifetimes enables plants to respond to environmental cues by redirecting their growth to exploit environmental opportunities, as when roots grow toward a water supply. We will return to plant–environment relationships in Chapter 28.
In Chapter 21 we saw how modern plants arose from aquatic ancestors, giving rise to simple land plants and then vascular plants. Despite their obvious differences in size and form, all vascular plants have essentially the same simple structural organization. This chapter describes the basic architecture of the largest group of vascular plants, the angiosperms (flowering plants), and shows how so much diversity (there are more than 250,000 species of angiosperms) can literally grow out of such a simple basic form. In this chapter we focus on the angiosperms’ three kinds of vegetative (nonsexual) organs: roots, stems, and leaves. We will cover reproductive organs in Chapter 27.
Plant organs are organized into two systems (FIGURE 24.1):
- The root system anchors the plant in place. Roots absorb water and dissolved minerals and store the products of photosynthesis. The extreme branching of roots and their high surface area-to-volume ratios allow them to absorb water and mineral nutrients from the soil efficiently.
- The shoot system consists of the stems, leaves, and flowers. The leaves are the chief organs of photosynthesis. The stems hold and orient the leaves to the sun and provide connections for the transport of materials between roots and leaves. The flowers are shoots modified for reproduction.
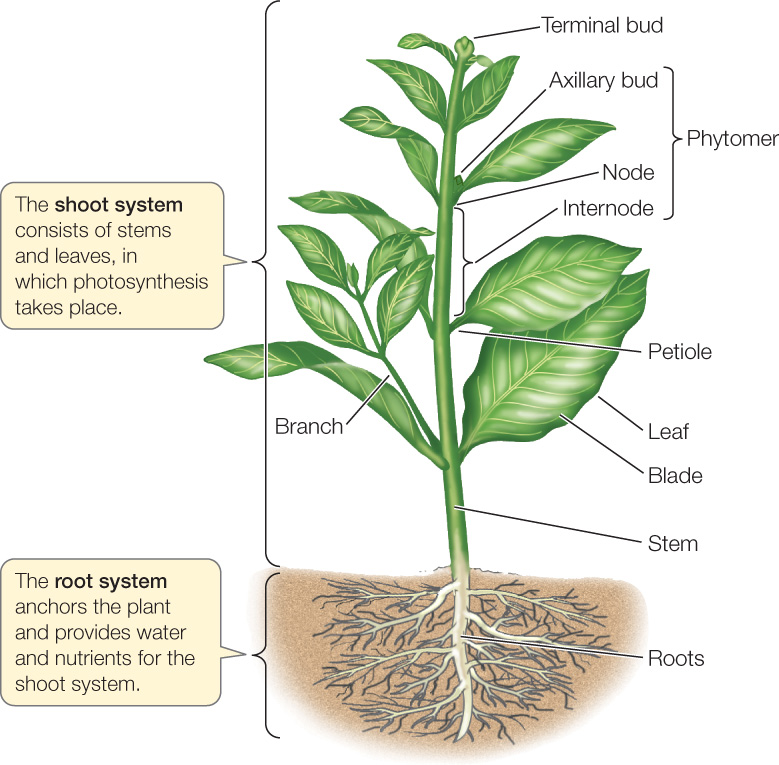
Shoots are composed of repeating modules called phytomers. Each phytomer consists of a node carrying one or more leaves; an internode, which is the interval of stem between two nodes; and one or more axillary buds, each of which forms in the angle where the leaf meets the stem. A bud is an undeveloped shoot that can produce another leaf, a phytomer, or a flower.
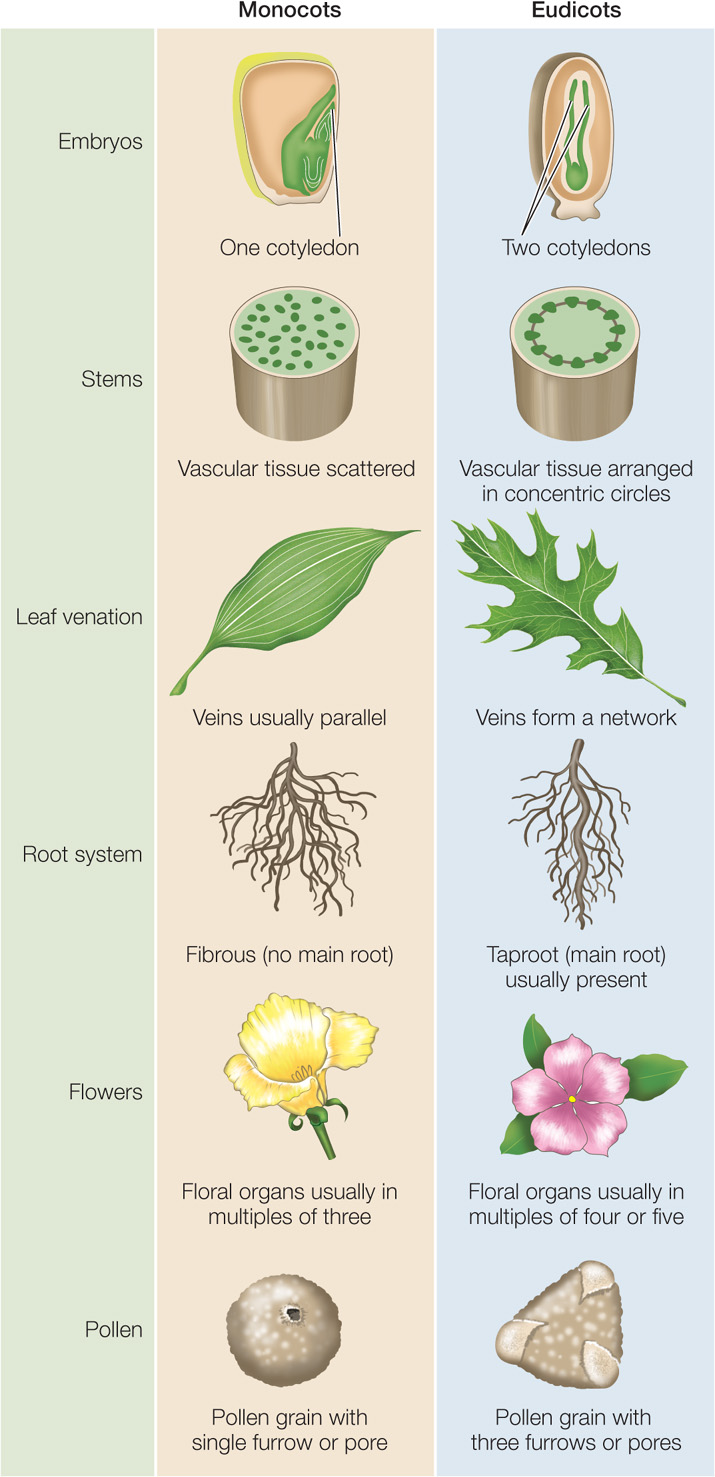
As noted in Chapter 21, most angiosperms belong to one of two major clades (see Figure 21.27). Monocots are generally narrow-leaved plants such as grasses, lilies, orchids, and palms. Eudicots are generally broad-leaved plants such as soybeans, roses, sunflowers, and maples. These two clades, which account for 97 percent of angiosperm species, differ in several important structural characteristics (FIGURE 24.2).
Plants develop differently than animals
The four processes that govern the development of all organisms, whether plant or animal, are determination (the commitment of an embryonic cell to its ultimate fate in the organism); differentiation (the specialization of a cell); morphogenesis (the organization and spatial distribution of cells into tissues and organs); and growth (increase in body size). In plants, these processes are influenced by three properties unique to plants: meristems, cell walls, and the totipotency of most cells.
LINK
The processes of development are described in Concept 14.1
Meristems
Plants grow throughout their lifetimes. Although adult humans, for example, have stem cells to replace tissues lost through damage or apoptosis, these cells do not contribute to further growth. In contrast, plants have meristems that are capable of producing new roots, stems, leaves, and flowers throughout the plant’s life, enabling the plant to continue growing.
Cell Walls
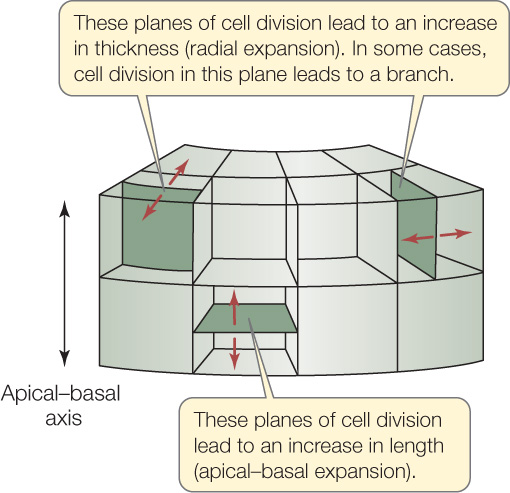
Plant cells are surrounded by a cell wall. This rigid extracellular matrix makes it impossible for cells to move from place to place as they do in animal development. Instead, plant morphogenesis occurs through alterations in the plane of cell division at cytokinesis, which in turn change the direction of cell expansion and thereby the direction in which a piece of tissue grows (FIGURE 24.3). Plant cytokinesis occurs along a cell plate laid down by Golgi vesicles (see Figure 7.7B). Unlike animal cells, in which the location of cytokinesis depends on the location of the middle of the mitotic spindle, the location of the plant cell plate is determined earlier—during interphase.
523
Totipotency
In animals, only early embryonic cells are totipotent, although some stem cell populations, such as those in bone marrow, are multipotent (capable of differentiating into a few different cell types). In contrast, in plants even differentiated cells are pluripotent, and most are totipotent (see Figure 14.3). This means a plant can readily repair damage wrought by the environment or herbivores.
LINK
See Concept 13.4 for the use of totipotency in plant biotechnology
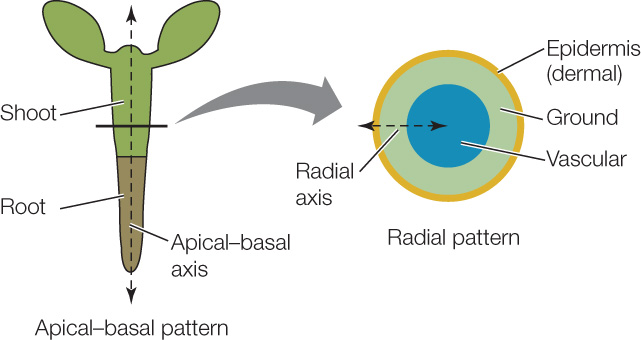
The plant body has an apical–basal axis and a radial axis
Two basic patterns are established early in plant embryogenesis (embryo formation) (FIGURE 24.4):
- The apical–basal axis: the arrangement of cells and tissues along the main axis from root to shoot
- The radial axis: the concentric (circular) arrangement of the tissue systems (which we will discuss in the next section) Both axes are best understood in developmental terms. We focus here on embryogenesis in the model eudicot Arabidopsis thaliana, in which the process has been intensively studied.
524
The first step in the formation of a plant embryo is a mitotic division of the zygote that gives rise to two daughter cells (FIGURE 24.5, STEP 1). An asymmetrical plane of cell division results in an uneven distribution of cytoplasm between these two cells, which face different fates. Signals in the smaller, apical (upper) daughter cell induce it to produce the embryo proper, while the other, larger daughter cell produces a supporting structure, the suspensor (FIGURE 24.5, STEP 2). This division not only establishes the apical–basal axis of the new plant but also determines its polarity (which end is the tip, or apex, and which is the base). A longer, thinner suspensor and a more spherical or globular embryo are distinguishable after just four mitotic divisions (see Figure 14.1).
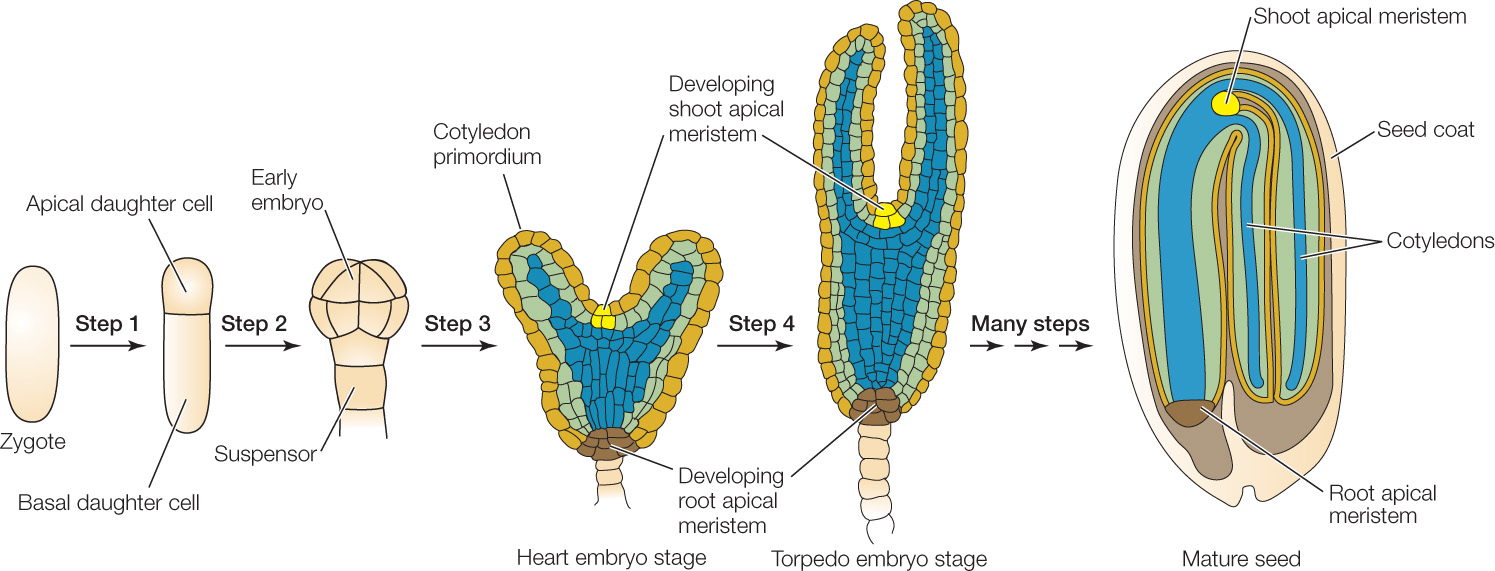
In eudicots, the initially globular embryo develops into the characteristic heart stage as the cotyledons (“seed leaves”) start to grow (FIGURE 24.5, STEP 3). Further elongation of the cotyledons and of the apical–basal axis of the embryo gives rise to the torpedo stage, during which some of the internal tissues begin to differentiate (FIGURE 24.5, STEP 4). Between the cotyledons is the shoot apical meristem; at the other end of the axis is the root apical meristem. These meristems contain undifferentiated cells that will divide to give rise to the shoot and root systems.
As shown in Figure 24.5, step 2, the plant embryo is first a sphere and later a cylinder. The root and stem retain a generally cylindrical shape throughout the plant’s life. You can see this most easily in the trunk of a tree. By the end of embryogenesis, the radial axis of the plant has been established. The embryonic plant contains three tissue systems, arranged concentrically, that will give rise to the tissues of the adult plant body.
The plant body is constructed from three tissue systems
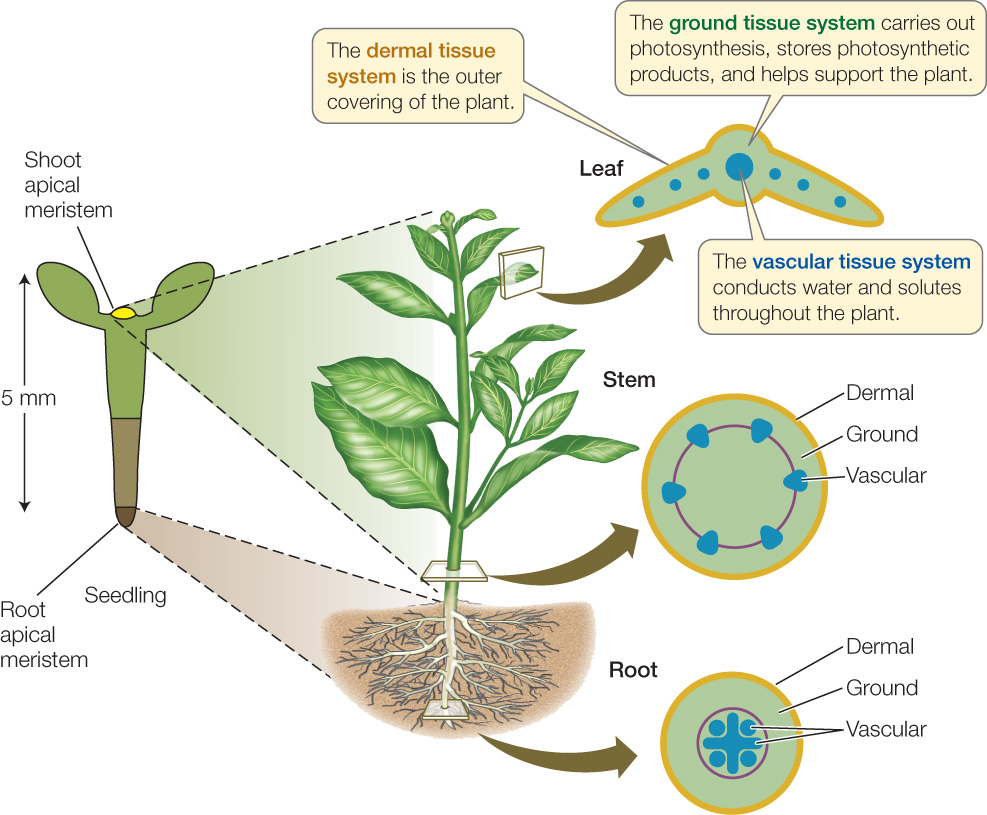
A tissue is an organized group of cells that have features in common and that work together as a structural and functional unit. In plants, tissues, in turn, are grouped into tissue systems. Despite their structural diversity, all vascular plants are constructed from three tissue systems: dermal, vascular, and ground. These three tissue systems are established during embryogenesis and ultimately extend throughout the plant body in a concentric arrangement (FIGURE 24.6). Each tissue system has distinct functions and is composed of different mixtures of cell types.
525
Dermal Tissue System
The dermal tissue system forms the epidermis, or outer covering, of a plant, which usually consists of a single cell layer. Epidermal cells are typically small. Some of them differentiate to form one of three specialized structures:
- Stomata, which are pores for gas exchange in leaves (see Figure 25.13)
- Trichomes, or leaf hairs, which provide protection against insects and damaging solar radiation (see Figure 28.7)
- Root hairs, which greatly increase root surface area, thus providing more surface for the uptake of water and mineral nutrients (see Figure 24.8)
Aboveground epidermal cells secrete a protective extracellular cuticle made of cutin (a polymer composed of long chains of fatty acids), a complex mixture of waxes, and cell wall polysaccharides. The cuticle limits water loss, reflects potentially damaging solar radiation, and serves as a barrier against pathogens.
As the stems and roots of woody plants expand, the epidermis is shed and is replaced by a dermal tissue called periderm. As we will see in Concept 24.3, this tissue includes an outer layer of cork that is impermeable to water and gases.
Ground Tissue System
Virtually all the tissue lying between dermal tissue and vascular tissue in both shoots and roots is part of the ground tissue system, which therefore makes up most of the plant body. Ground tissue contains three cell types.
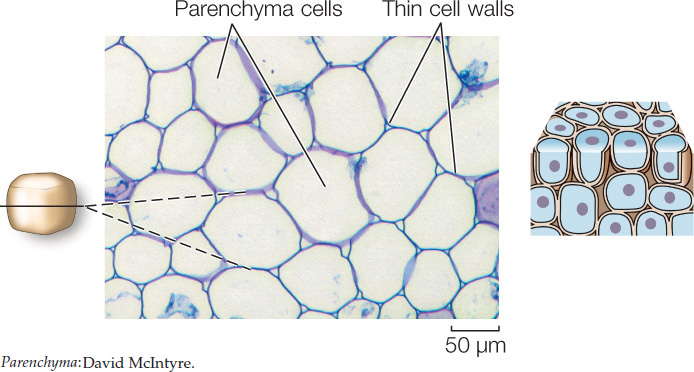
Parenchyma cells are the most abundant ground tissue cells. They have large vacuoles and relatively thin cell walls. They perform photosynthesis (in the shoot) and store protein (in seeds) and starch (in roots).
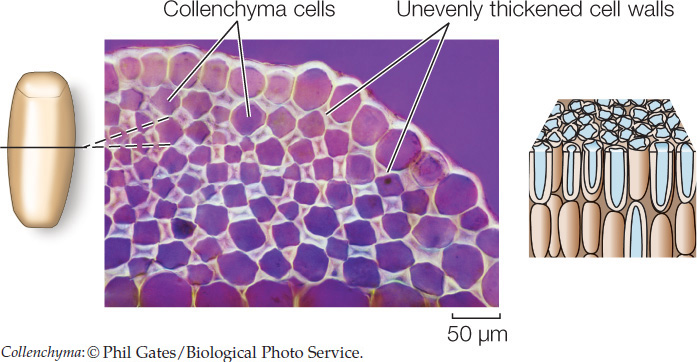
Collenchyma cells are elongated and have unevenly thickened primary cell walls. They provide support for growing tissues such as stems. The familiar “strings” in celery consist primarily of collenchyma cells.
Sclerenchyma cells have very thick secondary walls reinforced with the polyphenol polymer lignin. Most sclerenchyma cells undergo programmed cell death (apoptosis; see Concept 7.5), but their strong cell walls remain to provide support for the plant. There are two types of sclerenchyma cells: fibers and sclereids. Elongated fibers provide relatively rigid support to wood and other parts of the plant, within which they are often organized into bundles. When wood is used to make high-quality paper, the wood pulp must be treated extensively to break down and remove the lignin in the fibers.
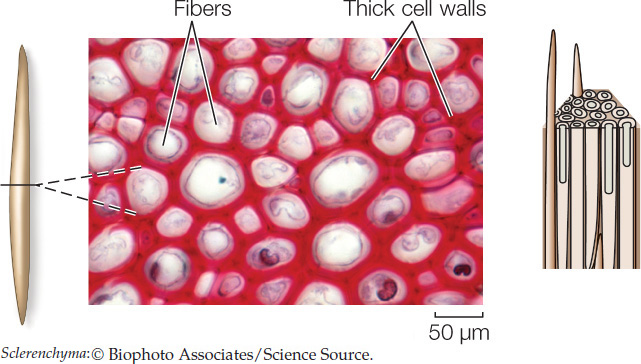
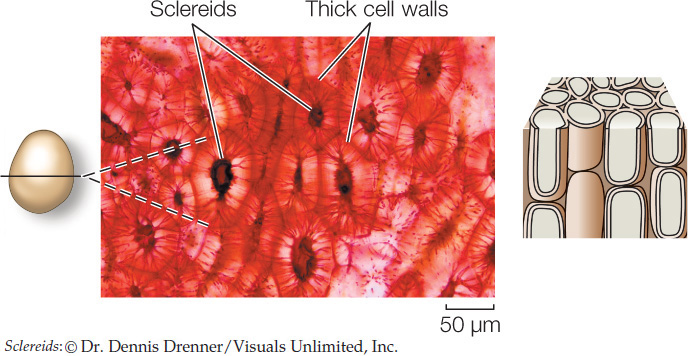
The bark of trees owes much of its mechanical strength to long fibers. Sclereids occur in various shapes and may pack together densely, as in a nut’s shell or in some seed coats.
526
Isolated clumps of sclereids, called stone cells, in pears and some other fruits give them their characteristic gritty texture.
Vascular Tissue System
The vascular tissue system is the plant’s plumbing or transport system—the distinguishing feature of vascular plants. Its two constituent tissues, the xylem and phloem, distribute materials throughout the plant.
LINK
The functions of xylem and phloem are described in
Concepts 25.3 and 25.4
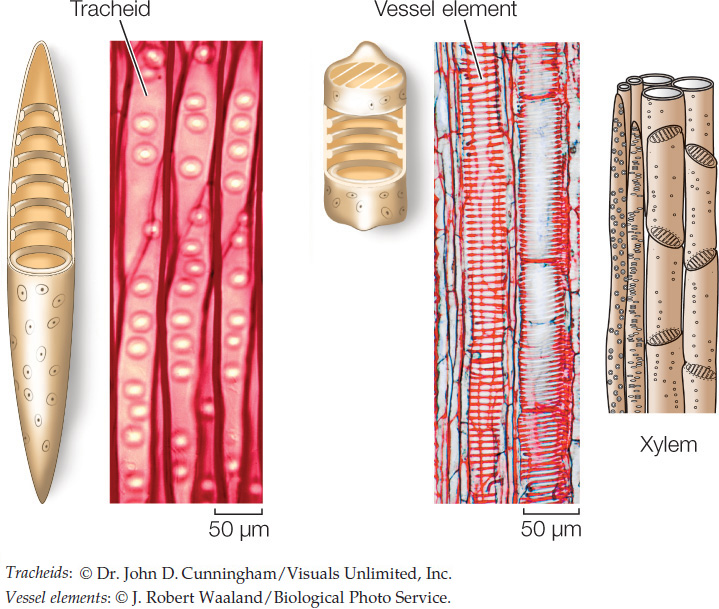
Xylem distributes water and mineral ions taken up by the roots to all the cells of the roots and shoots. It contains two types of conducting cells—tracheids and vessel elements—which, like most sclerenchyma cells, are dead when mature. Tracheids are spindle-shaped cells, with thinner regions in the cell wall called pits through which water can move with little resistance from one tracheid to its neighbors. Vessel elements are larger in diameter than tracheids. They meet end-to-end and partially break down their end walls, forming an open pipeline for water conduction. The xylem also contains living parenchyma cells that regulate the movement of ions into the conducting cells.
Phloem generally consists of living cells. It transports carbohydrates (primarily sugars) from sites where they are produced (called sources) to sites where they are used or stored (called sinks). Sources are often photosynthetic organs (such as leaves) but also include nonphotosynthetic storage organs (such as tubers and seeds) when they are drawn on to supply sugars to other parts of a plant. Sinks include growing tissues, roots, and developing flowers and fruit. The characteristic cells of the phloem are sieve tube elements, which, like vessel elements, meet end-to-end, forming sieve tubes. Instead of the ends of the cells breaking down, however, the sieve tube elements are connected by plasmodesmata (see Figure 4.7), which form a set of pores called a sieve plate. Although still alive, mature sieve tube elements have lost much of their cellular contents; adjacent, fully functional companion cells are connected to the sieve tube elements by plasmodesmata and perform many of the phloem’s metabolic functions.
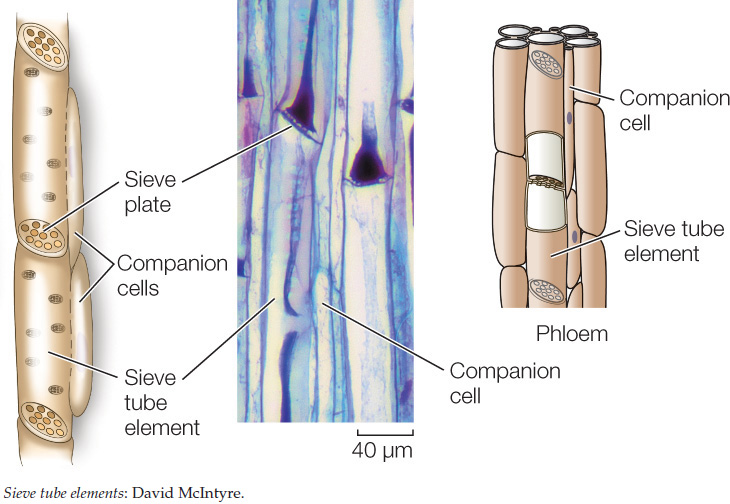
Xylem and phloem tissues may also contain fibers, which provide structural support.
CHECKpointCONCEPT24.1
- Make a table listing the three tissue systems of plants and their functions.
- What are the similarities and differences between plant and animal development?
- What are the similarities and differences between a root and a shoot?
- A mutant strain of Arabidopsis thaliana lacks apical–basal polarity in the embryo. What would you expect the phenotype of the plant to be?
We’ve discussed the basic body plan and tissue systems of plants, and how the apical–basal axis and radial axis arise during embryo development. Now let’s see how the cells and tissues we have just described allow the embryo to build an adult plant body.