CONCEPT28.1 Plants Have Constitutive and Induced Responses to Pathogens
Botanists know of dozens of diseases that can kill a wheat plant, each of them caused by a different pathogen, each of which in turn has many different genetic strains. Plant pathogens—which include fungi, bacteria, protists, and viruses—are part of nature, and for that reason alone they merit our study in biology. Because we humans depend on plants for our food, the stakes in our effort to understand plant pathology are especially high. That is why, just as medical schools have departments of pathology, universities in agricultural regions have departments of plant pathology.
Successful infection by a pathogen can have significant effects on a plant, reducing photosynthesis and causing massive cell and tissue death. Plants and pathogens have evolved together in a continuing “arms race”: pathogens have evolved mechanisms with which to attack plants, and plants have evolved mechanisms for defending themselves against those attacks. Like the responses of the human immune system, the responses by which plants fight off infection can be either constitutive—always present in the plant—or induced—produced in reaction to the presence of a pathogen.
Physical barriers form constitutive defenses
As with humans and their skin, a plant’s first line of defense is its outer surfaces, which can prevent the entry of pathogens. The parts of stems and leaves exposed to the outside environment are covered with cutin, suberin, and waxes. These substances not only prevent water loss by evaporation but can also prevent fungal spores and bacteria from entering the underlying tissues. The plant cell wall is also an important physical barrier to pathogens and can be strengthened further as an induced defense response (as we will see in the next section).
In addition to using physical barriers to prevent infection, plants use chemical warfare. Unlike animals, plants cannot flee from animals that eat them, so they make molecules that deter or destroy herbivores, as we will see in Concept 28.2. Some of these constitutively synthesized molecules also inhibit pathogens. For example, in plantains (Plantago), iridoid glycosides, which are glucose molecules covalently linked to short-chain fatty acids, inhibit the growth of fungal pathogens.
When constitutive defenses fail to deter a pathogen, plants initiate induced resistance mechanisms. As we discuss these mechanisms, refer to the overview in FIGURE 28.1.
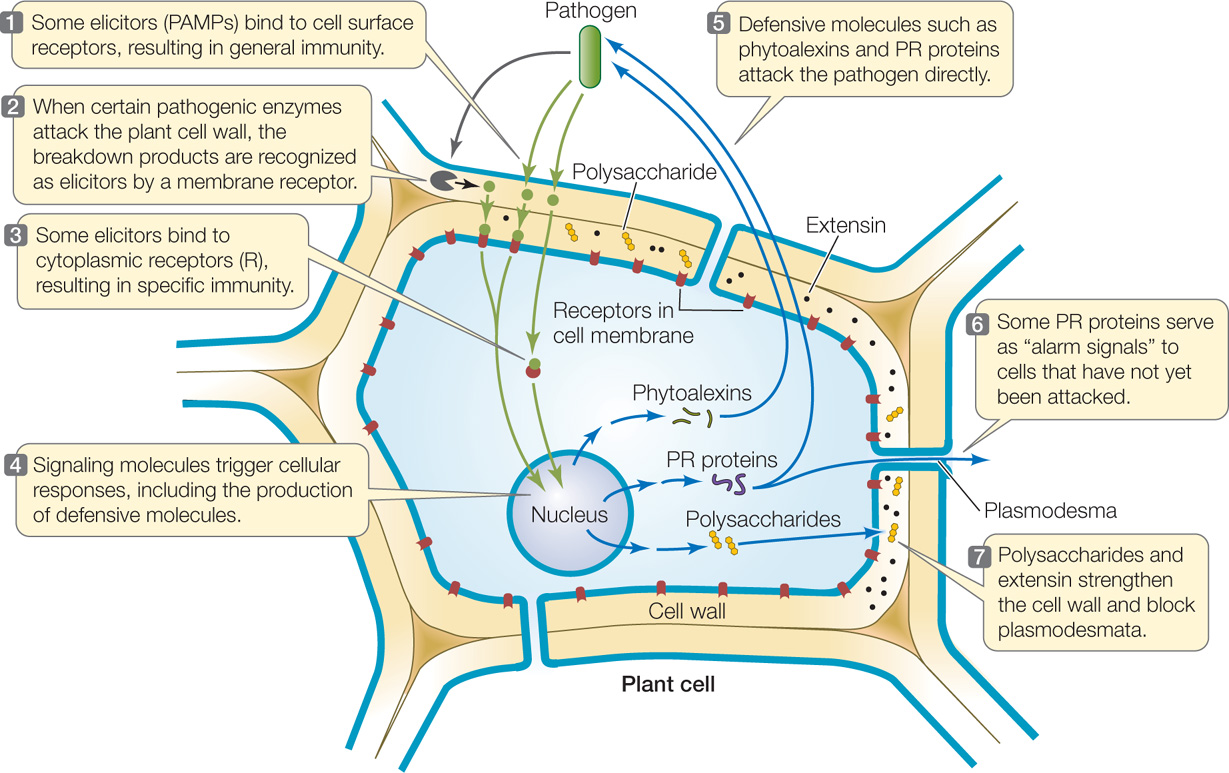
Induced responses can be general or specific
Plant pathogens cause the host plant to activate various chemical defense responses. A wide range of molecules called elicitors have been identified that trigger these defenses. These molecules vary from peptides made by bacteria to cell wall fragments from fungi. Elicitors can also be derived from fragments of plant cell wall components broken down by pathogens. Plants have receptors that recognize elicitors and initiate the induced defense responses.
The responses of plants to elicitors can be described in terms of the “plant immune system.” Two forms of immunity are recognized (see Figure 28.1):
591
- General immunity is triggered by general elicitors called pathogen associated molecular patterns (PAMPs). PAMPs are usually molecules that are produced by entire classes of pathogens, such as the protein flagellin (found in bacterial flagella) or the polysaccharide chitin (found in fungal cell walls). Thus general immunity is an overall response rather than a response that is triggered by a specific pathogen in a particular plant. PAMPs are recognized by transmembrane receptors called pattern recognition receptors, which activate signaling pathways that lead to general immunity.
- Specific immunity is triggered by specific elicitors called effectors. Effectors include a wide variety of specific pathogen-produced molecules that enter the plant cell. Once inside the cell, effectors bind to cytoplasmic receptors called R proteins that trigger the specific immunity response.
LINK
The human immune system, with its general (innate) and specific (adaptive) defenses, is described in Chapter 39
General and specific immunity both involve multiple responses
Many of the signaling pathways associated with general and specific immunity are the same, although the latter is specific for particular pathogens in particular plants, and is much stronger than general immunity. Both forms of immunity involve signaling pathways that are triggered by binding between the elicitors (PAMPs or effectors) and their receptors. These pathways lead to various responses:
- Formation of NO and reactive oxygen species: Receptor binding triggers the rapid production of nitric oxide (NO) and reactive oxygen species such as hydrogen peroxide (H2O2). These reactive molecules are toxic to some pathogens, and they are components of signal transduction pathways leading to local and systemic (plant-wide) defenses.
- Polymer deposition: Polymers such as lignin are deposited on the inside of the cell wall to strengthen the wall. These macromolecules may also block the plasmodesmata, limiting the ability of pathogens to spread from cell to cell through these channels (see Figure 28.1, step 7).
- Hormone signaling: Some pathways result in the production of plant hormones, including salicylic acid and jasmonic acid. We will describe the roles of these hormones in immunity later in the chapter.
- Changes in gene expression: Signal transduction cascades lead to changes in gene expression. The upregulated genes include pathogenesis-related (PR) genes and genes encoding the production of antimicrobial substances called phytoalexins.
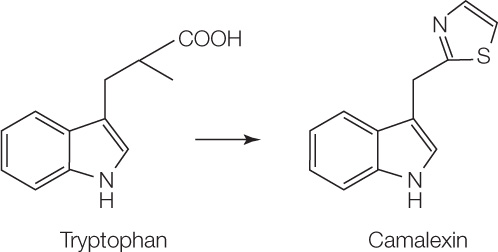
Phytoalexins
Phytoalexins are small molecules produced within hours of an infection by plant cells near the infection site. They are antibiotic-like molecules, toxic to pathogens. An example is camalexin, a phytoalexin made by the model organism Arabidopsis thaliana from the amino acid tryptophan:
Pathogenesis-Related Proteins
Plants produce several types of pathogenesis-related, or PR, proteins. Some are enzymes that break down the cell walls of pathogens. Chitinase, for example, is a PR protein that breaks down chitin, a distinctive component of fungal cell walls. In some cases the breakdown products of the pathogen’s cell walls serve as elicitors that trigger further defensive responses. Other PR proteins may serve as alarm signals to plant cells that have not yet been attacked, so they can initiate defensive responses of their own (see Figure 28.1, step 6). PR proteins are not rapid-response weapons; rather, they act more slowly, perhaps after other mechanisms have blunted the pathogen’s attack.
Plants have constitutive and induced responses to pathogens
Because you are a biology student, your nonscientist neighbors are confident that you know all about plants. A neighbor brings you a potted plant he bought 3 weeks ago from a nursery. The leaves have brown spots. He bought another plant of the same species from a different nursery 3 weeks ago, and that plant has no spots.
- You suspect that the spots are caused by either a disease or a nutritional deficiency. Given a large supply of the plants and soils involved, how would you distinguish between these possibilities?
- Assuming that the spots are caused by a fungal disease, how would you investigate at the molecular level the mechanism for formation of the brown leaf spots?
- You go to the nursery where the neighbor bought the infected plant and find several pots of plants with no brown spots on their leaves. What plant characteristics might account for the absence of spots?
592
Specific immunity is genetically determined
Pathogen genes that code for elicitors of specific immunity are called avirulence (Avr) genes; there are hundreds of such genes, and they vary among pathogen species and strains. When an elicitor enters a plant cell, it may encounter an R protein, a cytoplasmic receptor protein encoded by a resistance (R) gene. There are hundreds of R genes, each encoding a receptor that is specific for one or a few elicitors. If a plant receptor binds to an elicitor, a signal transduction pathway is set in motion that triggers the plant’s specific immunity. This type of resistance is called gene-for-gene resistance because it involves a recognition event between two specific molecules: one determined by an Avr gene in the pathogen and the other determined by an R gene in the plant (FIGURE 28.2).
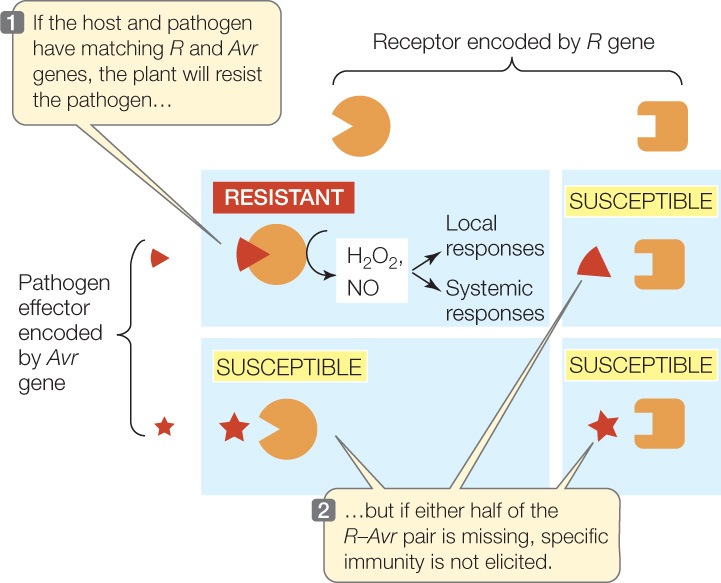
If the plant has no receptor to bind to the elicitor made by a particular species or strain of pathogen, the plant does not recognize the pathogen and does not turn on specific inducible defenses. The lack of a receptor makes the plant especially susceptible to that pathogen. There is an ongoing coevolutionary “arms race” between pathogens attempting to evade detection by plants and plants evolving ways to detect them. A major goal of plant breeders for the past 50 years has been to identify R and Avr genes and to breed new R genes into crops to make them more resistant to pathogens. This effort speeds up the arms race that occurs naturally.
Specific immunity usually leads to the hypersensitive response
As we described earlier, some induced defenses, such as polymer deposition, help prevent spread of the pathogen to the rest of the plant. Specific immunity often takes this a step further: cells around the site of infection undergo apoptosis (programmed cell death) in what is called the hypersensitive response. This prevents spread of the pathogen by depriving it of nutrients. Some of the cells also produce phytoalexins and other toxic chemicals before they die. Surrounding (still living) cells produce cell wall polysaccharides to seal off plasmodesmata and prevent the spread of the pathogen to healthy tissues. The dead tissue, called a necrotic lesion, contains and isolates what is left of the infection. The rest of the plant remains free of the infecting pathogen:

LINK
Apoptosis, or programmed cell death, is involved in many normal biological processes; see, for example, Concept 14.3. The cellular events of apoptosis are described in Concept 7.5
General and specific immunity can lead to systemic acquired resistance
The hypersensitive response contrasts with systemic acquired resistance, a general increase in the resistance of the entire plant to a wide range of pathogens. It is not localized and not limited to the pathogen that originally triggered it, and its effect may last as long as an entire growing season. This defensive response is initiated by salicylic acid:
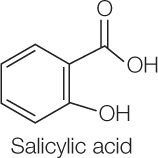
Salicylic acid production is triggered by receptor binding in both general and specific immunity; salicylic acid then acts as a signal to turn on various defense responses.
Systemic acquired resistance is accompanied by the synthesis of PR proteins. Infection in one part of a plant can lead to the export of salicylic acid to other parts, where it triggers the production of PR proteins ahead of the spread of infection. Infected plant parts also produce the closely related compound methyl salicylate (also known as oil of wintergreen). This volatile (readily forms a gas) substance travels to other plant parts through the air and may trigger the production of PR proteins in neighboring plants that have not yet been infected.
Another type of systemic acquired resistance is a more specific defense against viruses whose genomes are made of RNA. The plant uses its own enzymes to convert some of the single-stranded RNA of the invading virus into double-stranded RNA (dsRNA) and to chop that dsRNA into small pieces called small interfering RNAs (siRNAs). The siRNAs interact with another cellular component to degrade viral mRNAs, blocking viral replication. This phenomenon is an example of RNA interference (RNAi) (see Figure 13.9). The siRNAs spread rapidly through the plant by way of plasmodesmata, providing systemic resistance.
593
CHECKpointCONCEPT28.1
- Compare constitutive and induced responses to pathogens.
- Explain the inherited traits involved in both constitutive and induced responses to pathogens.
- If a group of plants is sprayed with oil of wintergreen, they all show resistance to a variety of pathogens. Explain how this occurs.
Pathogens are not the only foes of plants; plants must also defend themselves against herbivores, animals that eat plants. Because they are sessile, plants cannot run away from herbivores, but that does not mean they are defenseless against their would-be consumers, as we will see next.