CONCEPT45.5 Rapid Climate Change Affects Species and Communities
Climate change is not expected to affect all ecosystems in the same way because the changes are not distributed evenly around the globe or across the seasons. As we have seen, some parts of the planet are getting drier while others are getting wetter (see Figure 45.14), and some parts are warming much faster than others. In all cases, however, recent climate change has been far more rapid than what organisms have experienced in their evolutionary histories.
Rapid climate change presents ecological challenges
The life histories of organisms are shaped by their environment, as we saw in Concept 42.3. Thus organisms that live in seasonal environments have adaptations to ensure that crucial life events occur at favorable times of the year. Alpine plants, for example, start growing when the snow melts; seeds of desert plants germinate in response to certain combinations of soil temperature and moisture; changes in day length often trigger plant flowering, bird migration, and onset or cessation of insect dormancy. But climate change is altering the relative timing of many such environmental cues: in a warmer world, the day length that once signaled the start of good growing conditions may instead indicate the middle of the growing season. Although species’ responses can evolve by natural selection, the rate of such evolution may be too slow, or available genetic variation may be too small, to keep up with an environment that changes too rapidly or that continues to change directionally for too long.
Over the short term, we have evidence that many species are continuing to track environmental changes so that events in their life histories remain within appropriate periods. For example, biological events that mark the end of the winter season in temperate ecosystems—such as the leafing out of trees—have generally been taking place at earlier dates as the climate has warmed. In Britain, the first flowers of a sample of 385 plant species have appeared, on average, 4.5 days earlier per 1°C increase in average temperature. However, this is an average value for advances in flowering, and not all species have responded in the same way—some did not shift their time of first flowering, and some now flower later in the spring rather than earlier. Thus some species may not be responding adaptively to climate change at all. Even for those that do, it is unclear that they can continue to track climate change over the long term.
Changes in seasonal timing can disrupt interspecific interactions
Not all environmental cues respond to the same degree, or in the same direction, to climate change, so relationships among cues are shifting. Because different species make use of different cues, they may respond differently to climate change, as the different British plant species have done. This variation raises the possibility of mismatches in the timing of life history events among species that interact with one another.
930
Biogeochemical cycles affect global climate
The graphs below show the results of computer simulations of Earth’s climate between 1850 and 2000. The blue-shaded regions indicate the range of average global temperatures produced by running the simulations many times (encompassing 95% of the outputs from the replicate simulations). The actual observed temperatures are shown by the red lines. The three graphs represent three different sets of assumptions used in the simulations: (A) only natural factors were included, such as solar radiation input, aerosols from volcanic eruptions, and oscillations in oceanic circulation; (B) only anthropogenic (human-generated) factors were included, such as land clearing, soot deposition, and emissions of aerosols and greenhouse gases; and (C) both natural and anthropogenic factors were included. Average global temperatures are expressed as deviation from the average global temperature of the last part of the nineteenth century (indicated by 0).a Using these graphs, answer the following questions, and explain your reasoning.
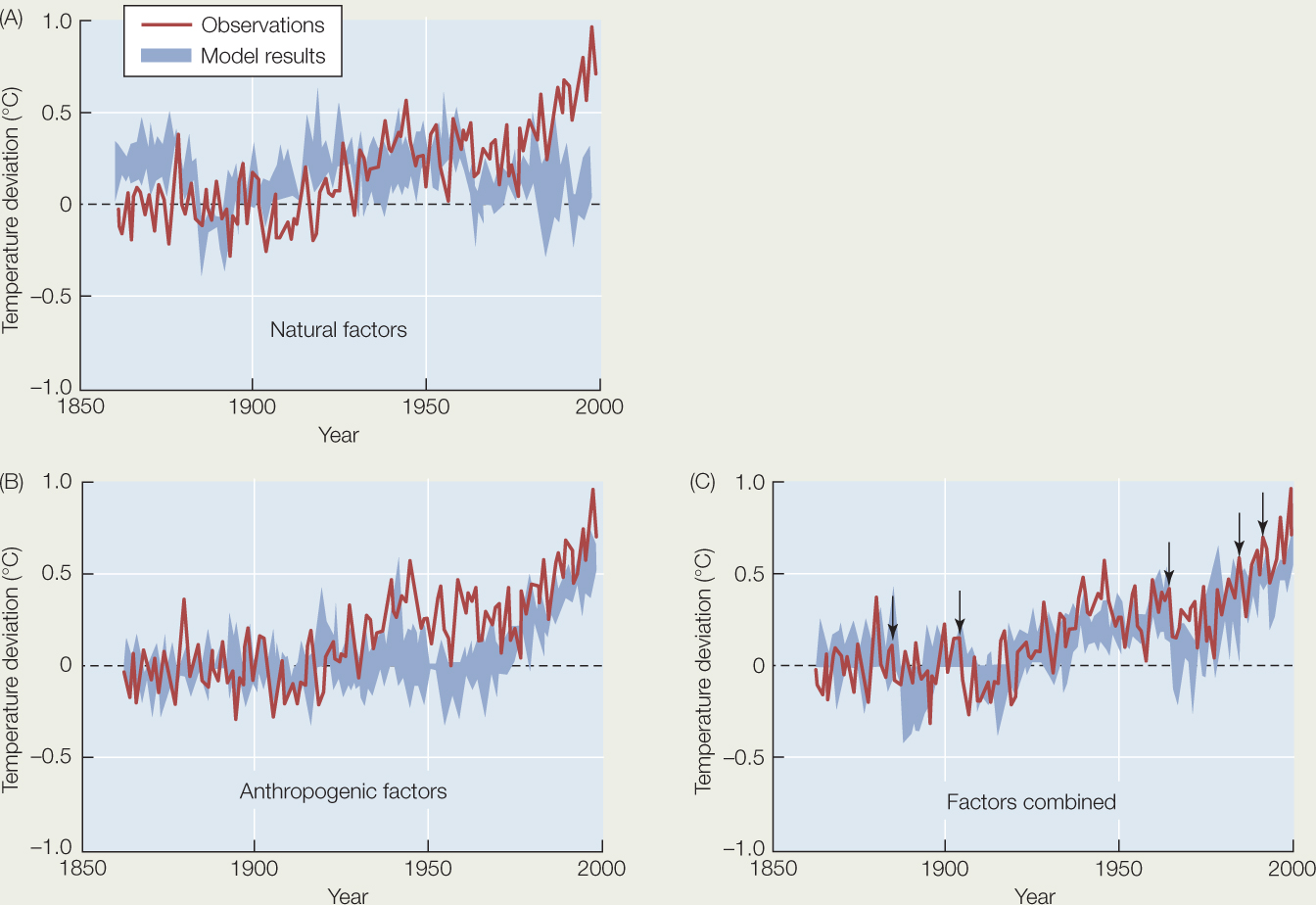
- What aspects of the observed trends in global temperature are not accounted for by natural factors?
- What aspects of the observed trends in global temperature are not accounted for by anthropogenic factors?
- Which set of factors best accounts for the observed global temperature trends?
- The arrows in panel (C) indicate five major volcanic eruptions. What effect did these natural events have on global temperatures?
Such mismatches are now becoming apparent in nature. In the Netherlands, for example, eggs of the winter moth (Operophtera brumata) hatch when temperatures warm after a fixed number of frosts, whereas the leaves of the oak trees on which the winter moth caterpillars feed (Quercus robur) emerge when warm days follow a certain number of cool days. The eggs have been hatching earlier, and oak leaves have been appearing earlier, in response to the earlier onset of warm temperatures in spring. The caterpillars do not use the same cues as the oaks, however, and they have been hatching too early and then starving. Great Tits (Parus major), which feed on winter moth caterpillars, are not nesting earlier because they use day length as the cue for egg laying (FIGURE 45.15). These birds now have less food for their nestlings not only because there are fewer caterpillars overall, but also because they are now feeding their nestlings after the peak in caterpillar abundance has passed. They are experiencing lower fecundity as a result.

Climate change can alter community composition by several mechanisms
If local populations cannot accommodate changing environmental conditions, they will go extinct, and their extinctions will change the species composition of their communities. As we saw in Figure 42.6, Sceloporus lizards in the Yucatán of Mexico are experiencing hotter temperatures during the spring breeding season, and this warming restricts the time during which they can forage. The lizards have gone locally extinct where they have insufficient feeding time to support reproduction. Extrapolating to lizards worldwide, researchers predict that continued global warming will place most lizard populations at risk of extinction by the year 2080. Losses of lizards may affect populations of their insect prey, of the plants those insects feed on, and of other animals that also eat the insects. As we saw in Concept 44.3, loss of diversity in communities can lead to overall loss of community function.
931
Altered relationships between environmental cues can also change communities even if species do not go locally extinct. In southwestern North America, the winter rainy season now arrives later in the fall, after soil temperatures have cooled. This shift favors plant species whose seeds use cool-season moisture as the cue to germinate, at the expense of species whose germination is cued by warm-season moisture. Thus the relative abundance of the cool-season species is increasing in these communities.
Climate-caused shifts in environmental gradients can also lead to the assembly of novel communities (see Concept 44.2). Some species have moved up mountains as their low-elevation habitats become too warm and higher-elevation habitats become more favorable. Others have moved toward higher latitudes for similar reasons; such poleward shifts have been documented for numerous taxonomic groups. Different species shift to different degrees or not at all (as is true of shifts in timing of life history events), so the mixture of species at any place changes. These distributional shifts also influence species interactions. Warmer temperatures in western North America, for example, are speeding up the life history of the mountain bark beetle (Dendroctonus ponderosae) and decreasing its winter mortality rate. As a result, beetle populations have increased and expanded, causing widespread tree mortality in pine and spruce forests from New Mexico to Alaska. Loss of the conifers has cascading effects on the consumers that depend on them, as well as on water runoff, wildfire frequency, and NPP.
Extreme weather events also have an impact
One of the predictions of climate models is that the frequency of extreme weather events will increase. Severe storms, flooding, droughts, heat waves, and other such events can elicit sudden shifts in species distributions and in community composition. For example, a brief but intense drought in the 1950s caused a ponderosa pine forest in northern New Mexico to shrink abruptly, and drought-adapted piñon–juniper woodland to expand, by more than 2 kilometers in less than 5 years. The new community boundary remained stable even after the drought ended. This is the most rapid shift in the boundary between two terrestrial plant communities so far documented at the scale of an entire landscape.
Such a response to an out-of-the-ordinary event might suggest that species are always able to change their geographic distributions rapidly in response to climate change. But as is true with shifts in timing, it remains virtually certain that many species will be left behind by continued shifts in environmental conditions, and that the number of species, their relative abundances, and their interactions will be altered in communities worldwide.
CHECKpointCONCEPT45.5
- Extrapolating from the British plant data, what change might you anticipate in average first flowering date by the middle of this century, when atmospheric CO2 is predicted to reach double its preindustrial concentration and the average temperature in Britain is expected to rise by an additional 3°C?
- How might changes in the seasonal timing of some events (such as the onset of warm weather at the end of winter) affect mutualistic interactions between plants and insect pollinators?
The changes that human activities are causing in Earth’s biogeochemical cycles and in its climate have serious ecological implications. What can we do to address them?