Competition can occur through exploitation or direct interference, or it may be apparent competition
Exploitative competition Competition in which individuals consume and drive down the abundance of a resource to a point that other individuals cannot persist.
Competition can be categorized in several ways. We have seen numerous examples of exploitative competition, in which individuals consume and drive down the abundance of a resource to a point that other individuals cannot persist. For example, rodents and ants that compete for seeds in the desert experience exploitative competition. Sometimes competitors do not immediately consume resources but defend them, a type of competition known as interference competition. While interference competition is a direct interaction between two individuals, exploitative competition is considered indirect competition because it operates through a resource. Finally, two species can appear to compete for a shared resource, but in fact experience a negative effect from apparent competition. In this section, we will explore the concepts of interference competition and apparent competition.
Interference competition When competitors do not immediately consume resources but defend them.
Interference Competition: Aggressive Interactions
Aggressive interactions are an effective form of interference competition that occurs between species of animals. For example, in the deserts of New Mexico, two species of ants—the long-legged ants (Novomessor cockerelli) and the red harvester ants (Pogonomyrmex barbatus)—compete for seeds and for any insects they can catch and subdue. However, the long-legged ants have a unique method of competing. Early in the morning, they emerge from their nests, find the nests of the red harvester ants, and plug the nest entrances with stones and soil. As you can see in Figure 16.14, when researchers examined the effects of this aggression, they found that harvester ants with plugged entrances required several hours to unplug their nests before they could begin foraging for seeds. This direct aggression by the long-legged ants allows them to forage in the morning with minimal competition from the harvester ants.
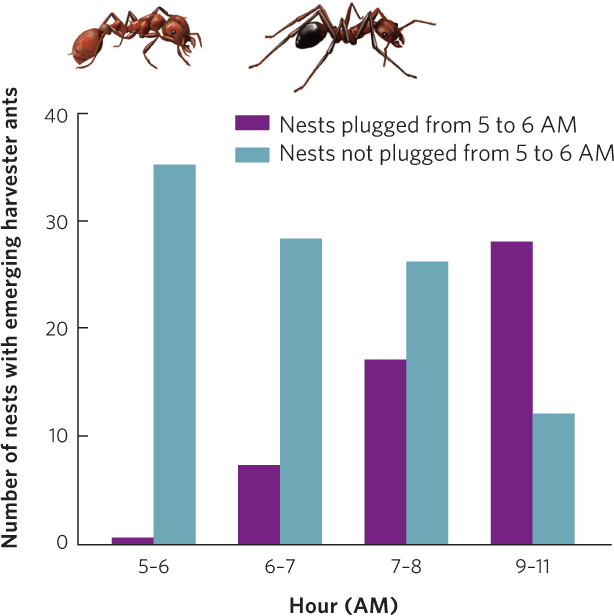
382
Interference Competition: Allelopathy
Allelopathy A type of interference that occurs when organisms use chemicals to harm their competitors.
Another type of interference, known as allelopathy, occurs when organisms use chemicals to harm their competitors. For example, black walnut trees (Juglans nigra) produce juglone, an aromatic organic compound that inhibits certain enzymes in other plants. The compound—found in the leaves, bark, roots, and seed husks of the black walnut tree—makes it difficult for most other species of plants to germinate and grow under a walnut tree.
Allelopathy can also be an effective strategy for invasive plants, which invade and dominate a community. The wetland plant known as the common reed (Phragmites australis) is found throughout the world (Figure 16.15). In North America, some genetic strains are native and other strains have been introduced from Europe. In locations such as the Great Lakes region of North America, the introduced strain is spreading rapidly and has displaced many other species of native wetland plants. For many years, researchers hypothesized that the common reed was successful at spreading due to allelopathy, but they lacked evidence. In 2007, researchers reported that the root of the common reed produced a chemical known as gallic acid, which is highly toxic to the roots of many other plants. By harming the roots of other species, the common reed is able to impair the growth of its interspecific competitors. The researchers also discovered that the chemicals produced by the introduced strains of the reed were much more lethal to some plants than those produced by the native strain, as shown in Figure 16.16. This result is consistent with the observation that the introduced strains of the reed are able to invade a wetland and spread much more rapidly than the native strains.

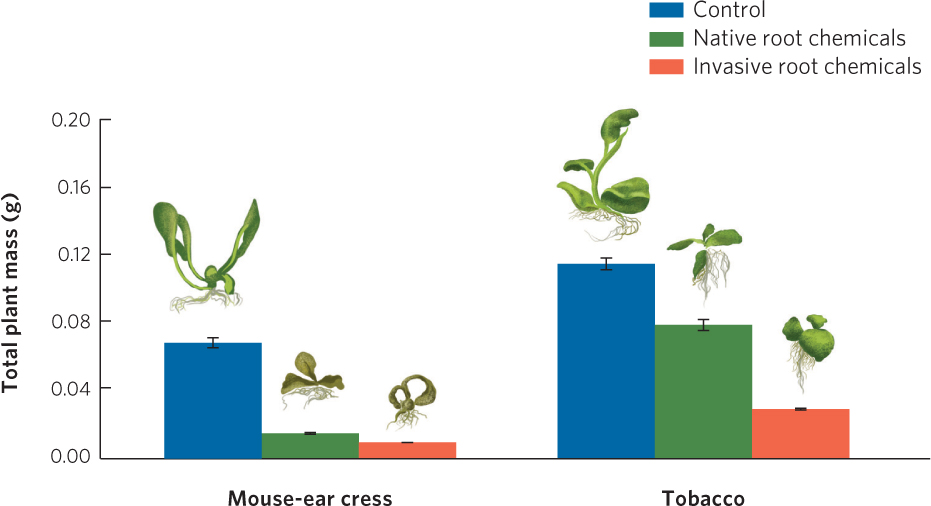
Allelopathy does not always take the form of toxic effects on interspecific competitors. In Australia, for example, researchers hypothesize that flammable oils in the leaves of some eucalyptus could be an adaptation that promotes frequent fires in the leaf litter of the forest floor, which kill seedlings of competitors.
Regardless of whether organisms use direct aggressive interactions or allelopathic chemicals, it is clear that interference competition can be an important mechanism in determining the abundance and distribution of species.
Apparent Competition
Apparent competition When two species have a negative effect on each other through an enemy—including a predator, parasite, or herbivore.
Throughout this chapter, we have defined competition as a negative interaction between two individuals for a limited resource. Sometimes, however, two species can share a resource and have negative effects on each other without competing for a resource. When two species have a negative effect on each other through an enemy—including a predator, parasite, or herbivore—we call it apparent competition. As its name suggests, apparent competition causes an outcome that looks like competition, but the underlying mechanism is different.
383
Apparent competition can be observed in a variety of communities. For example, the ring-necked pheasant and the gray partridge (Perdix perdix) live in many of the same habitats in the United Kingdom. For 50 years, a large decline in the number of gray partridges has been attributed to increased agriculture, although researchers suspected that a parasitic nematode (Heterakis gallinarum) might also play a role. Infected pheasants experience few harmful effects from the parasite, but infected partridges can suffer weight loss, reduced fecundity, and death.
To determine the susceptibility of each species to infection by parasites in the environment, researchers allowed both bird species to feed either in pens with parasite eggs scattered across the soil or in pens without the eggs. After 50 days of living in the pens without added parasite eggs, they found that both species contained low parasite infections, as shown in Figure 16.17. However, in the pens that contained added parasite eggs, the pheasants became infected at rates that were more than 20 times higher than the rate of infection in the partridges. Moreover, when both birds were fed the same number of nematode eggs over a 100-day period, the pheasants excreted over 80 times more nematode eggs than the partridges. Pheasants can carry a large number of parasites without being harmed and they excrete a large number of parasites that subsequently infect and harm partridges. The presence of pheasants is associated with partridges that gain less weight and have a low rate of survival. The relationship between the birds has the appearance of competitors for a shared resource, but the real cause is a shared parasite.
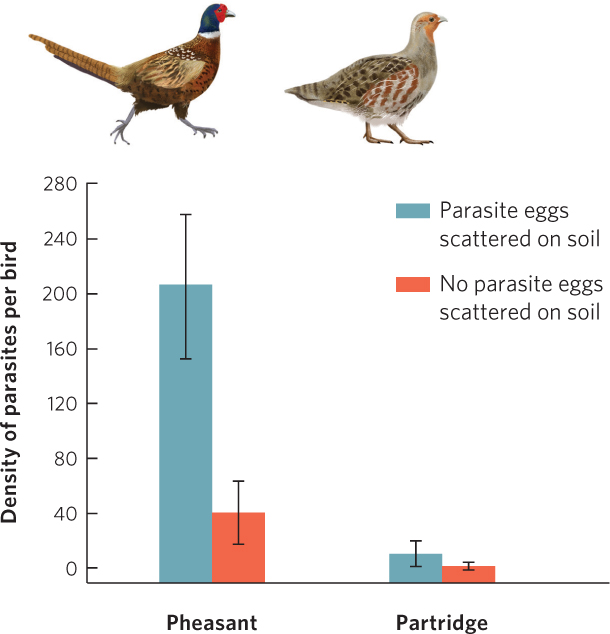
384
There are many examples of pathogens that cause apparent competition. For instance, when Europeans colonized other continents, particularly North and South America, they brought with them diseases such as smallpox. The Europeans could tolerate many of these diseases because they had a long evolutionary history of living with them, but the Native Americans lacked this evolutionary history and the introduced diseases devastated their populations. While the Europeans appeared to cause competition, the reduction of the Native American population occurred primarily because the Europeans brought new pathogens to the Western Hemisphere.
In some cases, apparent competition occurs not because of a shared enemy, but because one species facilitates the enemy of another species. For example, on the coast of California, several species of shrubs commonly have a zone of bare soil around them followed by a zone with a variety of grasses (Figure 16.18a). Researchers initially proposed that the bare soil was caused by allelopathic chemicals produced by the shrubs, which made sense because some of the shrub species were known to produce chemicals that inhibited the growth of other plants when the chemicals were at high concentrations in the soil. However, since not all of the shrub species produced these chemicals the allelopathy hypothesis was not well supported.
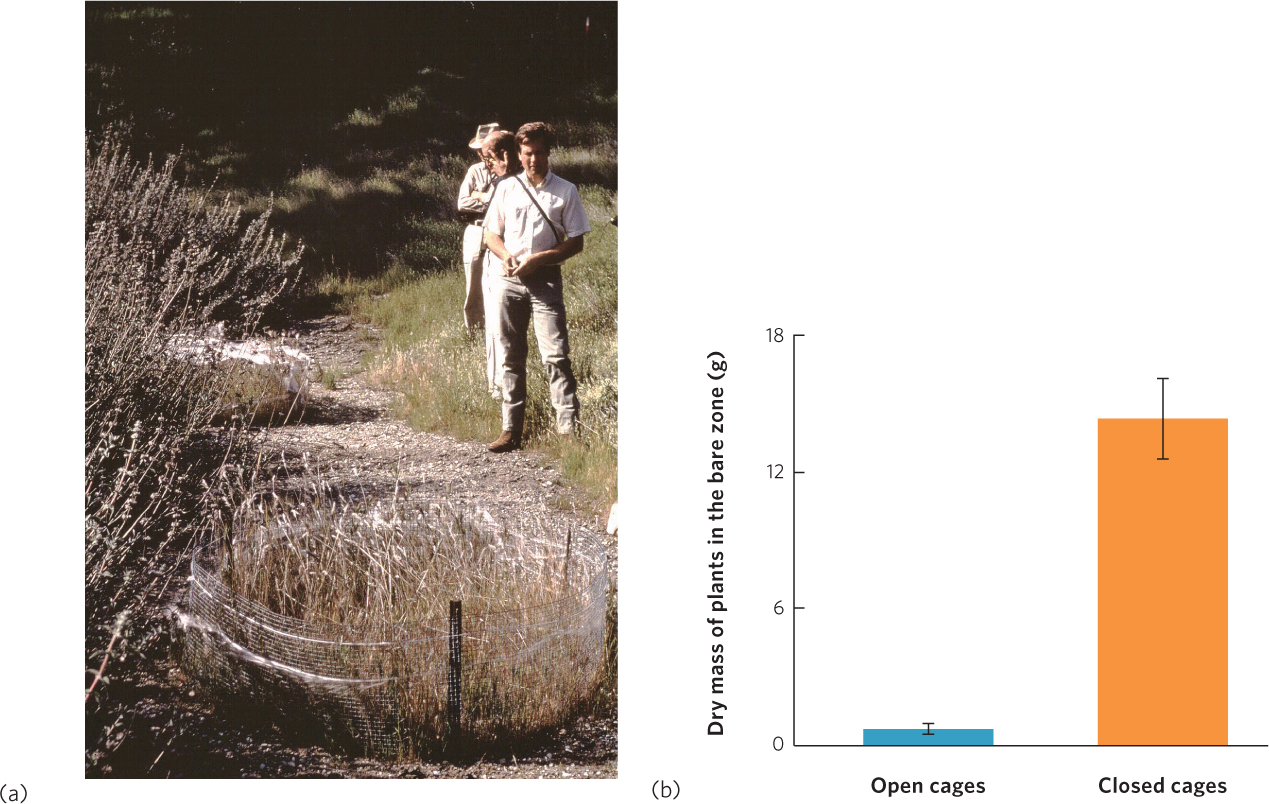
Further investigations found that what appeared to be the outcome of interference competition was not really competition at all. For example, when researchers put seeds on the bare zone and in the grass, they found that 86 percent of the seeds were consumed in the bare zone but only 12 percent were consumed in the grass. When they set out small mammal traps in both areas, they captured 28 mice in the bare zone but only 1 mouse in the grass. Finally, they placed two types of cages in the bare zone: open-sided cages that allowed mice to move in and out of the cages and closed cages that excluded rodents. One year later, there was still little vegetation in the open cages but nearly 20 times more vegetation in the closed cages, as shown in Figure 16.18b. Collectively, these results demonstrate that the bare zone, which looked to be the outcome of competition by allelopathy, actually occurred because the shrubs served as a safe haven for mice and other herbivores that primarily ate plants in close proximity to the shrubs.
385
ANALYZING ECOLOGY
Chi-square Tests
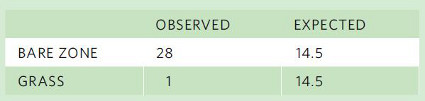
When the researchers in California were trying to determine the causes of the bare zone around coastal shrubs, they captured 28 mice in the bare zone near the shrubs and 1 mouse in the grass. Although this is a substantial difference, how do we know if this difference is statistically significant? In Chapter 14, we discussed the importance of testing hypotheses using a criterion of α < 0.05 for statistical significance. In Chapter 15 we discussed testing hypotheses using t-tests when we have estimates of mean values and standard deviations. Sometimes, however, our data do not consist of means and standard deviations, but rather of individuals that are counted, such as the number of mice in the bare zone versus those in the grass.
Chi-square test A statistical test that determines whether the number of observed events in different categories differs from an expected number of events, which is based on a particular hypothesis.
When our data consist of counts, we can test whether the data are significantly different from an expected distribution using a Chi-square test. A Chi-square test is a statistical test that determines whether the number of observed events in different categories differs from an expected number of events, which is based on a particular hypothesis. In the case of the mice, the researchers sampled both habitats equally and they captured a total of 29 mice from both areas. If the mice exhibited no habitat preference, we would have expected to find 14.5 mice in the bare zone and 14.5 mice in the grass. As a result, we want to statistically compare the observed distribution of mice—28 mice in the bare zone and 1 mouse in the grass—to an even distribution between the two habitats. Now that we know the observed distribution and the expected distribution for mice that exhibited no preference, we can create a small table and conduct the Chi-square test.
To calculate the Chi-square value, we use an equation that includes our observed (O) and expected (E) values to calculate the value of Chi-square (χ2).
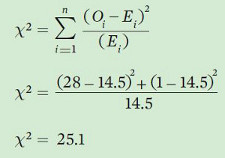
As was the case when we did a t-test in Chapter 15, we have to compare our calculated value to a table of values to determine if our observed distribution is significantly different from the expected distribution. The first step in that process is determining the degrees of freedom:
Degrees of freedom = (number of observed categories − 1) = (2 − 1) = 1
Now we can examine the Chi-square table provided in the Statistical Tables in the appendix. Using these tables, we can compare our calculated value (25.1) to the critical Chi-square value when we have 1 degree of freedom and we set α = 0.05. In this case, the critical Chi-square value is 3.841. Since our calculated value is greater than the critical value, we can conclude that the observed distribution of mice differed significantly from an equal distribution of mice between the two habitats.


Throughout this chapter, we have seen that competition can be a major force affecting the distribution and abundance of organisms. This competition can occur by exploitation or interference and it can occur among species both closely and distantly related. As we will see in “Ecology Today: Finding the Forest in the Ferns,” understanding the role of competition and how it interacts with disturbance, herbivory, and apparent competition is important for understanding how the natural world works, including biomes that provide humans with valuable natural resources.
386
ECOLOGY TODAY CONNECTING THE CONCEPTS
FINDING THE FOREST IN THE FERNS

In forests around the world, adult trees produce offspring that can become the next generation of trees. This next generation will be available to support the other species that live in the forest and will be available for such human uses as lumber and paper products. For a long time, it was assumed that the composition of trees in the next generation would be determined by the composition of seedlings produced by adult trees, the ability of each species to tolerate shade as a seedling, and the amount of shade that is present. For example, because seedlings of quaking aspen trees (Populus tremuloides) cannot tolerate shade from adult trees, they regenerate only when there are large openings in the forest that allow high amounts of sunlight. In contrast, seedlings of American beech trees are highly tolerant of shade and can grow in forests even under a dense canopy of leaves. While these processes are important determinants of the composition of the next generation of forest, they can be overwhelmed by processes that promote the dense growth of a single plant species that dominates the understory.
An excellent example of this phenomenon has occurred in the forests of central Pennsylvania. Historically, these forests had a variety of tree species in the upper canopy and a variety of younger trees in the understory, but over decades as the forests were logged, an abundance of plant growth occurred. In addition, top predators such as mountain lions and wolves were removed, which allowed a dramatic increase in the white-tailed deer population. Deer consumed large numbers of seedlings but not an unpalatable plant known as the hay-scented fern (Dennstaedtia punctilobula). Although the fern historically comprised less than 3 percent of the understory vegetation, today it is the dominant plant in more than one-third of Pennsylvania forests. Few seedlings can emerge out of the dense fern layer, so there are many large trees in the forest canopy but few young trees to serve as the next generation.
For many years, foresters hypothesized that ferns were causing competitive exclusion by casting high amounts of shade over the seeds and seedlings. It was thought that ferns prevented the germination and growth of the tree seeds and seedlings by consuming nearly all the light and making it a limiting resource. Competition as a mechanism between these distantly related species was completely consistent with the low number of tree seedlings that were emerging from the fern layer. However, an alternative explanation was that the fern layer provided an attractive refuge for rodents from predatory hawks and owls and that these rodents were consuming tree seeds and tree seedlings.
387
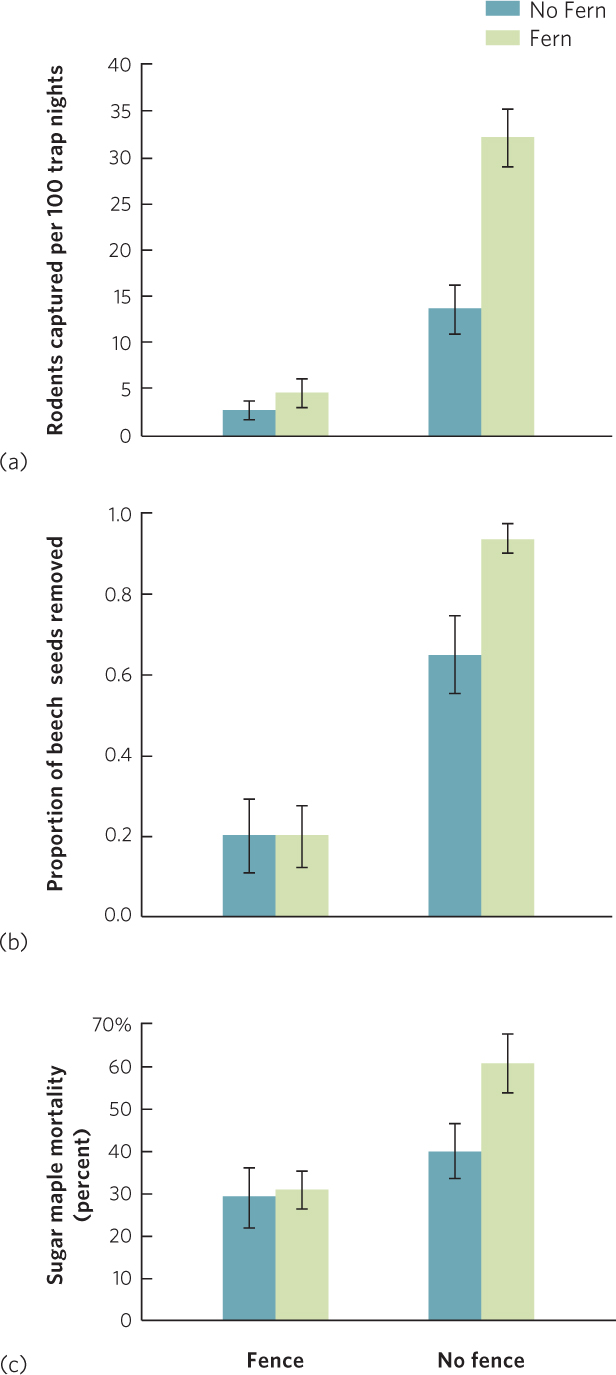
In 2008, researchers reported the results of an experiment to determine which of these two hypotheses was correct: whether the negative effect of the ferns was caused through competition for resources or through apparent competition by facilitating the rodents. They began by selecting plots of ferns within a forest, each with an area of 4 m2. For each plot, they either fenced out rodents or let rodents roam throughout the plot and they either removed the ferns or left them in place. After conducting the four different manipulations, the researchers began collecting data by first examining how many rodents were captured in each type of plot. The most common rodent species were deer mice, chipmunks, and red-backed voles (Clethrionomys gapperi). As you can see in part (a) of the graph, rodents rarely got inside the fenced plots, regardless of fern coverage. However, rodents were frequently captured in the unfenced plots and more than twice as many rodents were captured when the ferns remained in place. When the researchers added beech seeds to the plots, they found that only a small percentage of seeds were removed in the fenced plots—regardless of fern coverage—as shown in part (b) of the graph. However, a large percentage of seeds were removed in the unfenced plots, and even more if the ferns remained in place. Similar results were seen for many other species of seeds, including black cherry (Prunus serotina) and sugar maple (Acer saccharum). Finally, when the researchers set out seedlings of sugar maple, they observed low mortality in the fenced plots—regardless of whether ferns were removed—as shown in part (c) of the graph. In the unfenced plots, however, seedling mortality was higher, but only when ferns were present. These results confirmed that the primary effect of the ferns on tree seedlings was not as a competitor for light, but rather that they served as a refuge for rodents that eat the seeds and seedlings of beech, black cherry, and sugar maple. It was clearly a case of apparent competition.
A different outcome occurred when the researchers examined red maple (A. rubrum). Although seed predation occurred more often on red maple when ferns were present, many red maple seeds were present on the forest floor. In the unfenced plots, the presence of ferns caused fewer red maple seedlings to germinate. However, ferns had the same effect in the fenced plots, which indicated that the overall effect of ferns on red maple was not due to seed predation but to resource competition.
388
The results confirmed that, while fern effects on red maple were due to resource competition, the fern effects on American beech, black cherry, and sugar maple were due to apparent competition mediated through the fern’s protection of rodents. These results confirm that both competition and apparent competition can play substantial roles in determining the species that will compose the next generation of a forest and therefore determine the diversity of species and the future species that will be available for harvest.
SOURCES: Royo, A, and W. C. Carson. 2006. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Canadian Journal of Forest Research 36: 1345–1362.
Royo, A, and W. C. Carson. 2008. Direct and indirect effects of a dense understory on tree seedling recruitment in temperate forests: Habitat-mediated predation versus competition. Canadian Journal of Forest Research 38: 1634–1645.